MOB4 vs. MOB1: Decoding the Antagonistic Regulation of Hippo Signaling in Development and Disease
The Hippo signaling pathway is a central regulator of tissue homeostasis, organ size, and tumor suppression.
MOB4 vs. MOB1: Decoding the Antagonistic Regulation of Hippo Signaling in Development and Disease
Abstract
The Hippo signaling pathway is a central regulator of tissue homeostasis, organ size, and tumor suppression. While the canonical MST1/2-MOB1-LATS1/2 kinase cascade is well-established, the non-canonical regulation by the STRIPAK complex, particularly through its component MOB4, presents a fascinating counterpoint. This article synthesizes current research to explore the antagonistic relationship between MOB4 and MOB1. We detail their distinct structures, binding partners (MST4 for MOB4 versus MST1/2 for MOB1), and opposing biological outcomes—where the MOB1 complex acts as a tumor suppressor, the MST4-MOB4 complex can promote oncogenesis. Aimed at researchers and drug development professionals, this review covers foundational concepts, methodological approaches for studying these interactions, troubleshooting for experimental challenges, and validation strategies, concluding with the translational implications for targeting this regulatory axis in cancer and other diseases.
Core Components and Antagonistic Functions: MOB1 as Hippo Activator vs. MOB4 as STRIPAK Inhibitor
The Hippo pathway is an evolutionarily conserved signaling network that plays a fundamental role in controlling organ size, tissue homeostasis, and tumor suppression by coordinately regulating cell proliferation and apoptosis [1] [2]. At the heart of this pathway lies the canonical MST1/2-MOB1-LATS1/2 kinase cascade, a critical tumor suppressor axis that restricts the activity of the oncogenic transcriptional coactivators YAP (Yes-associated protein) and TAZ (transcriptional coactivator with PDZ-binding motif) [3] [2]. When the Hippo pathway is active, this core kinase cassette phosphorylates and inactivates YAP/TAZ, preventing their translocation to the nucleus and thus inhibiting the expression of genes that promote cell proliferation and survival [1] [3]. Dysregulation of this axis leads to aberrant activation of YAP/TAZ, contributing to uncontrolled tissue growth and neoplasia, highlighting its essential role as a guardian against tumor development [1] [3].
Table 1: Core Components of the Canonical Hippo Pathway
| Component | Drosophila Homolog | Function | Phenotype of Loss |
|---|---|---|---|
| MST1/2 | Hippo (Hpo) | Serine/threonine kinase that initiates the cascade | Tissue overgrowth, increased progenitor cells [1] |
| MOB1A/B | Mats (Mob as tumor suppressor) | Adaptor protein that activates LATS1/2 [4] | Uncontrolled tissue growth [1] |
| LATS1/2 | Warts (Wts) | Serine/threonine kinase that phosphorylates YAP/TAZ | Tissue overgrowth, expanded progenitor cells [1] |
| YAP/TAZ | Yorkie (Yki) | Transcriptional coactivators, pathway effectors | Nuclear translocation induces proliferation genes [1] |
| TEAD1-4 | Scalloped (Sd) | Transcription factors that bind YAP/TAZ | Mediates expression of growth-promoting genes [1] |
Molecular Architecture of the MST1/2-MOB1-LATS1/2 Axis
Stepwise Activation Mechanism
The canonical Hippo pathway operates through a precisely regulated, phosphorylation-dependent kinase cascade. The activation mechanism follows these sequential steps:
MST1/2 Activation: The kinase cascade can be initiated by TAO kinases (TAOK1/2/3), which phosphorylate the activation loop of MST1/2 (Thr183 for MST1 and Thr180 for MST2), or through MST1/2 autophosphorylation facilitated by dimerization via their C-terminal SARAH domains [1] [4]. Active MST1/2 complexed with the scaffold protein SAV1 then phosphorylates both LATS1/2 and MOB1A/B [1] [2].
MOB1 as Central Signal Transducer: MOB1 plays a dual role as both a scaffold and an allosteric activator. MST1/2 phosphorylates MOB1 at its N-terminal residues (T12 and T35 in humans), which relieves MOB1 autoinhibition and enables its binding to LATS1/2 [5] [4]. Recent structural studies reveal that phospho-MOB1 undergoes conformational activation and binds to the N-terminal regulatory domain of LATS1, creating an active kinase complex [5] [4].
LATS1/2 Activation and YAP/TAZ Phosphorylation: The MOB1-LATS1/2 interaction facilitates LATS1/2 phosphorylation by MST1/2 at conserved hydrophobic motifs (T1079 for LATS1 and T1041 for LATS2), triggering LATS1/2 autophosphorylation and full activation [1] [4]. Active LATS1/2 then phosphorylates YAP/TAZ, leading to their cytoplasmic sequestration through 14-3-3 binding and subsequent ubiquitin-mediated degradation [1] [3].
Structural Basis of Kinase Activation
The molecular interactions within the MST1/2-MOB1-LATS1/2 axis are governed by precise structural mechanisms:
MST1/2-MOB1 Interaction: Active MST1/2 autophosphorylates multiple residues in the flexible linker region between its kinase and SARAH domains, creating phospho-docking motifs that bind MOB1. This binding converts MOB1 from an autoinhibited, closed conformation to an active, open conformation capable of LATS1/2 binding [4].
MOB1-LATS1/2 Interface: Phosphorylated MOB1 binds to the N-terminal regulatory domain of LATS1/2 through complementary electrostatic surfaces. The crystal structure of the MOB1/LATS1 complex reveals that MOB1's acidic surface interacts with conserved basic residues in LATS1's N-terminal domain [5] [4].
Allosteric Activation of LATS1/2: The phosphorylated N-terminal tail of MOB1 and LATS1's phospho-hydrophobic motif act synergistically to allosterically promote LATS1 autophosphorylation in its activation loop, leading to full kinase activation [4].
Experimental Approaches for Studying the Canonical Axis
Key Methodologies and Reagents
Table 2: Essential Experimental Protocols for Investigating the MST1/2-MOB1-LATS1/2 Axis
| Methodology | Key Reagents/Assays | Experimental Readout | Significant Findings |
|---|---|---|---|
| Co-immunoprecipitation & Pulldown Assays | FLAG/HA-tagged constructs, λ protein phosphatase, kinase-inactive mutants (e.g., MST4-K53R) [6] | Protein-protein interactions, phosphorylation-dependent binding | MST4-MOB4 complex disrupts MST1-MOB1 assembly [6] |
| X-ray Crystallography & Structural Analysis | Truncation mutants (e.g., MST4 linker regions 316-335), MOB1 variants with selective loss-of-interaction mutations [5] [6] | Atomic-level complex structures, interface residues | Identification of Asp63 as key MOB1 residue for LATS1 binding [5] |
| Kinase Activity Assays | Phospho-specific antibodies (pMob1 T35), quantitative immunoblotting, recombinant purified proteins [4] | Phosphorylation status, kinase activation | MOB1 phosphorylation at T12/T35 essential for LATS1 binding [4] |
| Functional Cellular Assays | YAP/TAZ localization, TEAD reporter assays, gene knockout/knockdown (MST1/2, MOB1, LATS1/2) [5] [2] | YAP/TAZ nuclear/cytoplasmic ratio, target gene expression, cell proliferation | MOB1-Warts binding essential for tumor suppression [5] |
| In Vivo Genetic Studies | Drosophila models, tissue-specific knockout mice, patient tumor samples [5] [2] | Tissue growth, tumor development, survival correlation | MST4-MOB4 upregulation in pancreatic cancer with poor prognosis [6] |
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Hippo Pathway Investigation
| Reagent/Category | Specific Examples | Function/Application |
|---|---|---|
| Expression Constructs | MOB1 selective loss-of-interaction variants (impaired MST1/2 or LATS1/2 binding) [5] | Decouple functional interactions in the kinase cascade |
| Kinase Modulators | Kinase-inactive mutants (MST2D146N, MST4-K53R) [4] [6] | Study phosphorylation-dependent complex assembly |
| Phospho-Specific Antibodies | pMOB1 (T12, T35), pLATS1 (T1079), pYAP antibodies [4] | Monitor pathway activation status in cells and tissues |
| Structural Biology Tools | Truncated proteins (MST2 linker residues 376-400, MOB1 33-216) [5] [4] | Determine atomic structures of complex interfaces |
| Cell Line Models | HEK293A, PANC-1, cancer cells with pathway mutations [1] [6] | Investigate pathway regulation in different contexts |
The STRIPAK-MOB4 Regulatory Counterpoint
MOB4 as a Non-Canonical Regulator
While MOB1 functions as a core activator in the canonical Hippo pathway, MOB4 represents a critical regulatory counterpoint that integrates Hippo signaling with broader cellular networks. MOB4, initially identified as phocein, is a highly conserved non-catalytic adaptor protein that serves as a core component of the STRIPAK (Striatin-interacting phosphatase and kinase) complex [7] [6]. Unlike MOB1, which activates LATS1/2 kinases, MOB4 lacks the capacity for stable binding to NDR/LATS kinases and instead associates with Hippo and Hippo-like kinases as part of the STRIPAK phosphatase complex [8] [7].
The MST4-MOB4 complex exhibits an overall structure similar to the MST1-MOB1 complex but has evolved to serve opposing biological functions. While the MST1-MOB1 complex suppresses oncogenic YAP activity, the MST4-MOB4 complex promotes growth and migration in cancer cells such as PANC-1 pancreatic cancer cells [6]. This functional divergence occurs despite structural conservation, highlighting the evolutionary plasticity of MST-MOB partnerships.
Mechanism of Pathway Antagonism
MOB4 antagonizes canonical Hippo signaling through two primary mechanisms:
Competitive Disruption: Due to divergent evolution of key interface residues, MST4 and MOB4 can disrupt the assembly of the tumor-suppressive MST1-MOB1 complex through alternative pairing. This disruption increases YAP activity and promotes oncogenic signaling [6].
STRIPAK-Mediated Dephosphorylation: As part of the STRIPAK complex, MOB4 contributes to the dephosphorylation and inactivation of MST kinases by recruiting the serine/threonine protein phosphatase 2A (PP2A). This phosphatase activity directly counteracts the kinase activity of the canonical Hippo cascade [7].
The expression patterns of these opposing complexes further highlight their yin-yang relationship: in pancreatic cancer, MST4 and MOB4 expression levels are elevated and positively correlated with each other, while MST1 expression is down-regulated [6]. This inverse relationship demonstrates the pathological relevance of this regulatory mechanism in human cancer.
Functional Consequences and Therapeutic Implications
Biological Outcomes of Pathway Dysregulation
The precise regulation of the MST1/2-MOB1-LATS1/2 axis determines critical cellular decisions and tissue-level outcomes:
Tumor Suppression: Genetic studies in Drosophila and mammalian models demonstrate that intact MST1/2-MOB1-LATS1/2 signaling is essential for proper development and tissue growth control. Deletion of any core component results in tissue overgrowth, expanded progenitor cells, and tumorigenesis [1] [5]. Specifically, the MOB1-Warts (LATS1/2) interaction is indispensable for tumor suppression, while stable MOB1-Hippo (MST1/2) binding appears dispensable for some developmental functions [5].
Therapeutic Targeting: The Hippo pathway's dysregulation in multiple cancer types has stimulated drug development efforts targeting this axis. Compounds such as Verteporfin and C19 show promise as cancer therapeutics by modulating YAP/TAZ activity [3]. Additionally, the opposing functions of MOB1 and MOB4 provide multiple potential intervention points—either enhancing tumor-suppressive canonical signaling or inhibiting the oncogenic STRIPAK-MOB4 axis [7] [6].
Integration with Other Pathways: The MST1/2-MOB1-LATS1/2 axis intersects with multiple oncogenic signaling networks, including TGFβ, Notch, Wnt, and EGFR pathways [3]. These connections enable the Hippo pathway to function as a signaling hub that coordinates diverse inputs to determine cellular fate decisions in development and disease.
The canonical MST1/2-MOB1-LATS1/2 axis represents a critical tumor-suppressive signaling module that governs tissue growth and homeostasis through a precisely regulated kinase cascade. The molecular dissection of this pathway has revealed sophisticated structural mechanisms involving phosphorylation-dependent conformational changes, dynamic scaffolding, and allosteric kinase activation. The emerging understanding of MOB4 as a counter-regulatory component through its integration into the STRIPAK complex adds an additional layer of complexity to Hippo pathway regulation, demonstrating how evolutionary diversification of MST-MOB partnerships can create opposing functional outputs. Future research focusing on the therapeutic manipulation of these interactions holds significant promise for cancer treatment and regenerative medicine applications.
The Hippo signaling pathway is an evolutionarily conserved mechanism crucial for regulating tissue homeostasis, organ size, and tumor suppression [7]. The canonical Hippo cascade involves a kinase cascade where MST1/2 kinases phosphorylate and activate the LATS/MOB1 complex, which subsequently phosphorylates and inactivates the oncogenic transcriptional coactivators YAP/TAZ, thereby inhibiting expression of proliferative and anti-apoptotic genes [7]. While MOB1 has been well-established as a core component activating LATS kinases within this pathway, recent research has unveiled a non-canonical regulatory axis centered on MOB4 and the STRIPAK complex that antagonizes Hippo signaling [8] [7].
The STRIPAK (Striatin-Interacting Phosphatase and Kinase) complex represents a large multisubunit PP2A-containing assembly that integrates diverse cellular signals [9]. As a kinase-phosphatase scaffold, STRIPAK contains both catalytic and structural subunits that position it as a key upstream modulator of Hippo pathway components [10] [11]. MOB4, initially identified as a neural interactor of striatins and alternatively named phocein, serves as an essential adaptor within this complex [7] [9]. Emerging evidence indicates that MOB4, through its incorporation into STRIPAK, regulates critical cellular processes including proliferation, migration, and neural development while functioning as a potent negative regulator of canonical Hippo signaling [6] [7].
Comparative Biology of MOB1 and MOB4
Structural and Functional Divergence
MOB1 and MOB4 belong to the highly conserved Mob family of adaptor proteins that function as critical interaction hubs without catalytic activity [8] [7]. Despite sharing a similar core structural fold consisting of a four-helix bundle, these proteins have diverged significantly in their interaction networks and biological functions.
Table 1: Comparative Properties of MOB1 and MOB4
| Property | MOB1 | MOB4 |
|---|---|---|
| Primary Complex | Hippo signaling core | STRIPAK complex |
| Binding Partners | MST1/2, LATS1/2 [6] | STRN3, STRIP1, PP2A, MST4 [9] |
| Hippo Pathway Role | Activator (Tumor suppressor) [7] | Negative regulator (Oncogenic) [6] |
| Cellular Localization | Cytoplasmic, nuclear [6] | Somato-dendritic, Golgi apparatus, dendritic spines [6] [7] |
| Biological Functions | Growth restriction, apoptosis promotion [7] | Cell proliferation, migration, neural development [6] [7] |
| Expression in Cancer | Often downregulated [6] | Frequently overexpressed [6] [7] |
The functional divergence between MOB1 and MOB4 stems from evolutionary changes in key interface residues that dictate specific binding partnerships [6]. While MOB1 preferentially associates with MST1/2 and LATS1/2 kinases to propagate Hippo signaling, MOB4 has lost the ability to bind NDR kinases like LATS but has gained specific affinity for components of the STRIPAK complex, particularly STRN3 [8] [9].
Molecular Mechanisms of Hippo Pathway Regulation
The opposing functions of MOB1 and MOB4 in Hippo signaling create a yin-yang regulatory relationship that fine-tunes YAP/TAZ activity in response to diverse cellular cues.
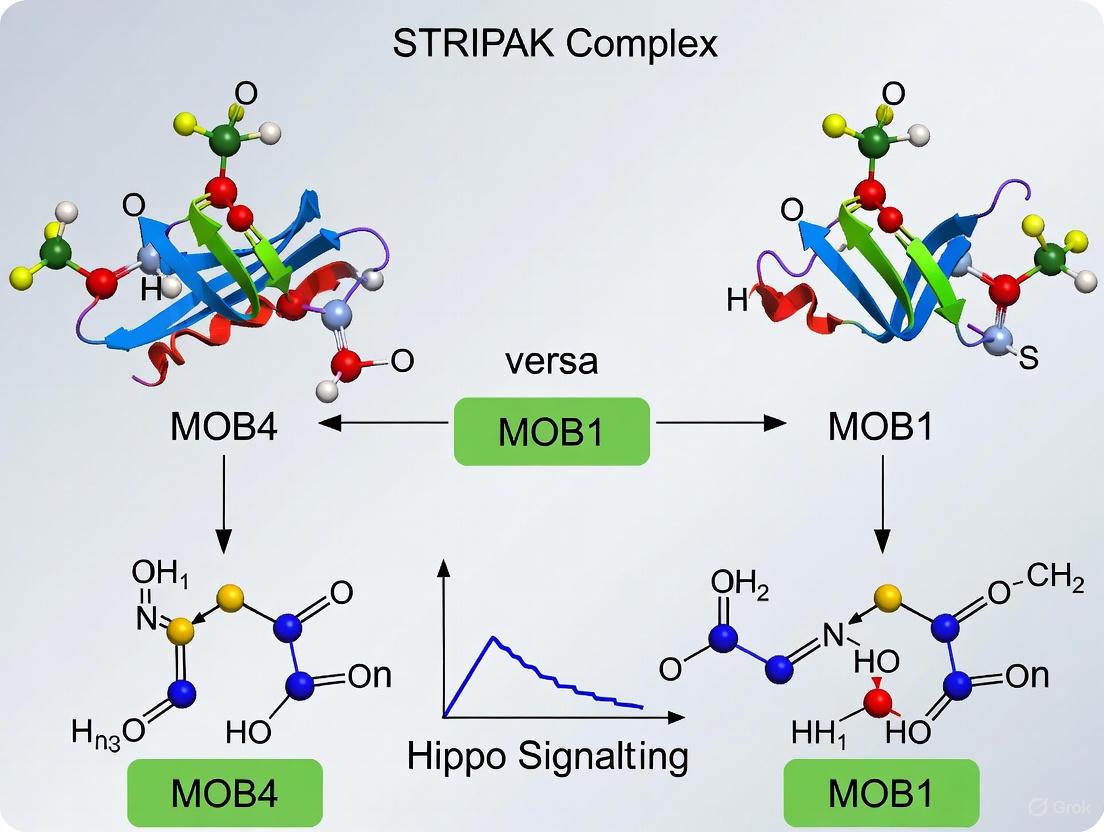
Figure 1: Regulatory networks of MOB1 and MOB4 in Hippo signaling. MOB1 (green) activates the canonical tumor-suppressive Hippo pathway, while MOB4 (red) integrated within STRIPAK inhibits MST1/2 to promote YAP/TAZ-mediated proliferation.
The canonical MOB1-dependent pathway initiates when MST1/2 kinases phosphorylate and activate MOB1, which subsequently binds and fully activates LATS1/2 kinases. Activated LATS1/2 then phosphorylate YAP/TAZ, leading to their cytoplasmic retention and proteasomal degradation [7]. In contrast, the non-canonical MOB4-STRIPAK axis disrupts this tumor-suppressive signaling through multiple mechanisms: (1) direct dephosphorylation and inactivation of MST1/2 kinases by the PP2A phosphatase within STRIPAK, (2) competitive binding wherein MST4-MOB4 complexes disrupt MST1-MOB1 assembly through alternative pairing, and (3) contextual regulation by cellular cues such as cell density [6] [11].
The STRIPAK Complex: Architecture and Assembly
Structural Organization of STRIPAK
The STRIPAK complex represents a non-canonical PP2A holoenzyme with a precisely defined architecture that distinguishes it from traditional trimeric PP2A complexes. Cryo-EM structural analysis has revealed that the core human STRIPAK complex comprises PP2AA, PP2AC, STRN3, STRIP1, and MOB4 with a striking 4:1:1:1:1 stoichiometry [9].
Table 2: Core Components of the STRIPAK Complex
| Component | Role in Complex | Functional Domains | Key Interactions |
|---|---|---|---|
| STRN3 (SG2NA) | Scaffold organizer | Coiled-coil, WD40 domain [10] | Forms homotetramer; binds PP2AA, STRIP1, MOB4 [9] |
| PP2AA/C | Catalytic phosphatase core | HEAT repeats (A), phosphatase domain (C) [9] | STRN3 coiled-coil (via PP2AA) [11] |
| STRIP1 | Central stabilizer | Unknown structure | Links PP2AC, STRN3, and MOB4 [9] |
| MOB4 | Adaptor protein | Mob/Phocein domain [8] | Binds STRIP1 and STRN3 WD40 domain [9] |
| CCM3 | Kinase recruiter | FAT domain, HBRCT domain [9] | Recruits GCKIII kinases (MST3/4, STK25) [9] |
The assembly mechanism centers on the STRN3 coiled-coil tetramer, which forms an elongated rod-like structure that serves as a structural platform for organizing other components [9]. This tetrameric arrangement creates a substantial scaffold with potential for integrating multiple signaling inputs, explaining the complex's role as a central signaling hub.
Dynamic Assembly and Regulation
STRIPAK complex assembly demonstrates context-dependent dynamics that enable responsive regulation of Hippo signaling. Research has revealed a "two-arm" model for STRIPAK organization, with PP2AA/C-bound STRN3 serving as an organizing center that recruits MST kinases through two distinct arms: one consisting of STRIP1 and the other comprising SIKE1-SLMAP [11].
Figure 2: "Two-arm" assembly model of the STRIPAK complex. STRN3 organizes the complex by simultaneously engaging STRIP1-MOB4 (Arm 1) and SIKE1-SLMAP (Arm 2) to recruit and regulate MST kinases.
This architecture exhibits signal-responsive dynamics – decreased cell density triggers dissociation of the STRIP1 arm from STRIPAK, loosening MST kinases from PP2A-mediated regulation [11]. This dynamic assembly allows STRIPAK to function as a cellular sensor that modulates Hippo signaling output in response to environmental cues.
Experimental Evidence and Methodologies
Key Experimental Findings
Multiple experimental approaches have demonstrated the functional opposition between MOB1 and MOB4 in Hippo signaling and their relevance in disease contexts, particularly cancer:
Pancreatic Cancer Studies: In PANC-1 pancreatic cancer cells, the MST4-MOB4 complex promoted growth and migration, contrasting with the tumor-suppressive effects of the MST1-MOB1 complex [6]. Expression analysis revealed elevated MST4 and MOB4 levels in pancreatic cancer tissues that positively correlated with each other, while MST1 expression was downregulated [6]. Crucially, the MST4-MOB4 complex increased YAP activity by disrupting MST1-MOB1 complex assembly through alternative pairing, highlighting a competitive interaction mechanism [6].
Neurological Development Research: Schwann-cell-specific ablation of STRN3 caused defects in lamellipodia formation and radial sorting during peripheral nervous system development [12]. Both Rac1 and striatin deletion in Schwann cells disrupted YAP/TAZ activation and expression of their target genes, particularly extracellular matrix receptors [12]. This established a functional connection between STRIPAK components, cytoskeletal reorganization, and Hippo pathway regulation in neural development.
Structural Biology Insights: Crystallography studies defined the phosphorylation-dependent interaction between MST4 and MOB4, with MST4 autophosphorylation at Thr-327/328 creating a critical binding interface for MOB4 [6]. Despite overall structural resemblance to the MST1-MOB1 complex, evolutionary divergence at key residue positions prevents cross-pairing and enables functional competition [6].
Essential Methodologies for MOB4-STRIPAK Research
Table 3: Key Experimental Protocols for Investigating MOB4-STRIPAK Function
| Method | Application | Key Technical Details | Representative Findings |
|---|---|---|---|
| Co-immunoprecipitation (Co-IP) | Detect protein-protein interactions | Endogenous vs. overexpression approaches; λ phosphatase treatment to test phosphorylation-dependence [6] | Confirmed MST4-MOB4 interaction is phosphorylation-dependent [6] |
| Pulldown Assays with Recombinant Proteins | Characterize direct interactions | MBP-tagged baits; kinase-active vs. kinase-dead mutants; mapping interaction domains [6] [11] | Identified MST4 linker region (aa 316-335) as MOB4-binding site [6] |
| Bio-layer Interferometry (BLI) | Quantify binding affinity | Dose-dependent measurements; determination of dissociation constants (Kd) [6] | Measured MST4-MOB4 Kd = 1.67 μM [6] |
| Cryo-EM Structural Analysis | Determine complex architecture | single-particle analysis; focused refinement; biGBac expression system for complex production [9] | Revealed STRN3 tetrameric organization in STRIPAK core [9] |
| Gene Knockdown/ Knockout | Functional characterization | shRNA in cell lines; conditional knockout in specific tissues [12] [13] | Schwann cell STRN3 ablation disrupts YAP/TAZ signaling [12] |
The Scientist's Toolkit: Essential Research Reagents
Table 4: Key Research Reagents for MOB4-STRIPAK Investigations
| Reagent/Cell Line | Specific Application | Function/Utility |
|---|---|---|
| PANC-1 Cell Line | Pancreatic cancer models | Assess oncogenic role of MST4-MOB4 complex [6] |
| HEK293FT Cell Line | Protein interaction studies | High transfection efficiency for Co-IP and complex purification [6] |
| λ Protein Phosphatase | Phosphorylation-dependence tests | Dephosphorylate proteins to confirm phosphorylation-dependent interactions [6] |
| STRN3 shRNAs | Functional knockdown studies | Target striatin family members to elucidate STRIPAK components [13] |
| MBP-Tagging System | Protein purification/pulldowns | Produce recombinant bait proteins for interaction mapping [6] [11] |
| Anti-pYAP Antibodies | Hippo pathway readout | Detect YAP S127 phosphorylation as indicator of pathway activity [13] |
Discussion and Research Perspectives
Therapeutic Implications
The opposing functions of MOB1 and MOB4 in Hippo signaling, coupled with their differential expression in cancers, highlight their potential as therapeutic targets. MOB4 overexpression in tumors may serve as a biomarker for YAP activation and represent a vulnerability that could be exploited therapeutically. The MST4-MOB4 interface, with its defined structural requirements, offers a potential target for small molecules that could disrupt this oncogenic pairing while preserving tumor-suppressive MST1-MOB1 interactions [6].
In neurological contexts, the role of MOB4-STRIPAK in Schwann cell development and neurite outgrowth suggests implications for peripheral neuropathies and nerve regeneration strategies [12] [7]. The enrichment of MOB4 in dendritic spines and its role in synaptic development further positions it as a potential modulator of neuronal plasticity and related disorders [7].
Unresolved Questions and Future Directions
Despite recent advances, several fundamental questions remain unanswered. The precise mechanisms governing dynamic assembly of STRIPAK subcomplexes in response to different cellular signals require further elucidation. The extent to which MOB4 functions independently of STRIPAK in certain contexts remains unexplored. Additionally, the regulatory hierarchy between different MST kinases (MST1/2 vs. MST3/4) in competing for MOB binding partners needs clarification.
Future research should prioritize developing conditional knockout models for tissue-specific analysis of MOB4 function, designing selective inhibitors targeting the MST4-MOB4 interface, and employing advanced proteomic approaches to identify context-dependent STRIPAK interactors. These investigations will further illuminate the complex regulatory networks orchestrated by MOB4 and the STRIPAK complex in health and disease.
MOB4, through its integration into the STRIPAK complex, represents a crucial non-canonical regulator that antagonizes the tumor-suppressive Hippo signaling pathway. The functional antagonism between MOB1 and MOB4 creates a delicate balance that fine-tunes YAP/TAZ activity in response to cellular context and environmental cues. Understanding the molecular details of this regulatory axis provides not only fundamental insights into cell signaling architecture but also potential therapeutic avenues for cancers and other pathologies driven by Hippo pathway dysregulation. As research continues to unravel the complexities of MOB4-STRIPAK biology, this emerging field promises to expand our understanding of cellular signaling integration in development, homeostasis, and disease.
Within the intricate landscape of intracellular signaling, the highly conserved Mob family proteins function as critical adaptors, directing kinase activity and pathway output. This guide provides a structural and functional comparison between two key family members: MOB1, a core component of the canonical Hippo tumor suppressor pathway, and MOB4, an integral member of the STRIPAK complex (Striatin-interacting phosphatase and kinase) that often antagonizes Hippo signaling [7] [14]. The Hippo pathway controls organ size and suppresses tumorigenesis by phosphorylating and inactivating the oncogenic co-activators YAP/TAZ [2]. Conversely, the STRIPAK complex, a large macromolecular assembly containing a phosphatase (PP2A), kinases, and scaffolding proteins, negatively regulates Hippo and other growth-restrictive pathways [15] [16]. The competition between MOB1 and MOB4 for binding to MST kinases represents a crucial regulatory node [6]. Understanding their distinct phospho-recognition mechanisms and binding interfaces is therefore essential for researchers dissecting cell proliferation, organ size control, and cancer development.
Structural Comparison of MOB1 and MOB4
MOB1 and MOB4 share a conserved core structural fold despite their divergent functions, belonging to a larger family of eukaryotic adaptor proteins that lack catalytic activity [14].
Table 1: Core Structural and Functional Attributes of MOB1 and MOB4
| Feature | MOB1 | B4 |
|---|---|---|
| Protein Family | Mob family, Class I [14] | Mob family, Class IV (Phocein) [7] [14] |
| Canonical Function | Activator of LATS1/2 in Hippo pathway [2] | Core component of STRIPAK complex [7] |
| Conserved Mob Fold | Yes, globular fold with four alpha-helix bundle core [14] | Yes, high structural homology to MOB1, including N-terminal region [7] |
| Key Pathway Role | Tumor suppressor [6] | Oncogenic promoter (context-dependent) [6] |
Comparative Analysis of Phospho-Recognition and Binding Interfaces
A critical distinction between MOB1 and MOB4 lies in their specific binding partners and the phosphorylation-dependent mechanisms governing these interactions.
Table 2: Comparative Binding Interfaces and Phospho-Recognition Mechanisms
| Characteristic | MOB1 | MOB4 |
|---|---|---|
| Primary Kinase Partner | MST1/2 (Hippo pathway kinases) [2] | MST4 (STE20-like kinase) [6] |
| Interaction Dependence | Phosphorylation-dependent [6] | Phosphorylation-dependent [6] |
| Critical Kinase Domain for Binding | MST1 linker region [6] | MST4 linker region (amino acids 316-335) [6] |
| Key Phospho-sites on Kinase Partner | Not fully detailed in results | MST4 autophosphorylation at Thr-327/Thr-328 [6] |
| Effect of Kinase-Inactive Mutant | Prevents complex formation (inferred) | MST4-K53R mutant cannot bind MOB4 [6] |
| Structural Complex | MST1-MOB1 complex [6] | MST4-MOB4 complex; resembles MST1-MOB1 structure [6] |
| Biological Outcome of Complex Formation | Activates LATS1/2, suppresses YAP, inhibits growth [6] [2] | Disrupts MST1-MOB1, activates YAP, promotes proliferation/migration [6] |
| Binding Affinity (Kd) | Not provided in results | 1.67 µM (MST4-MOB4 interaction) [6] |
The overall structure of the MST4-MOB4 complex closely resembles that of the canonical MST1-MOB1 complex, indicating evolutionary conservation of the binding mode [6]. However, despite this structural mimicry, they exert opposite biological functions in pathways like pancreatic cancer. The MST4-MOB4 complex promotes cell growth and migration, whereas the MST1-MOB1 complex suppresses it [6]. This functional antagonism is achieved through alternative pairing; MST4 and MOB4 can disrupt the assembly of the tumor-suppressive MST1-MOB1 complex, thereby increasing YAP oncogenic activity [6].
Experimental Data and Key Methodologies
The comparative insights between MOB1 and MOB4 are derived from robust structural biology and biochemical techniques.
Key Experimental Protocols for Characterizing Interactions
The following methodologies are essential for delineating the MOB-kinase interactions.
Co-immunoprecipitation (Co-IP) and Pulldown Assays: Used to confirm direct physical interactions in vivo and in vitro. For example, FLAG-tagged MST4 successfully pulls down HA-tagged MOB4 in HEK293FT cells, and endogenous co-IP confirms this association in other cell lines [6]. Pulldown assays with purified recombinant MBP-tagged MST4 and MOB4 protein demonstrate a direct interaction, excluding the need for other cellular factors [6].
Phospho-dependency Analysis: Treatment with λ protein phosphatase (λPP) is used to dephosphorylate proteins. Studies show λPP treatment markedly reduces the MST4-MOB4 interaction, while subsequent incubation with ATP/MgCl₂ (allowing autophosphorylation) restores it [6]. The use of a kinase-inactive mutant (MST4-K53R), which cannot bind MOB4, further confirms the phosphorylation-dependence of the complex [6].
Mapping Critical Binding Regions: A series of truncation mutants of the MST4 linker region are generated as MBP-fusion proteins. Following phosphorylation by wild-type MST4, these mutants are used in pulldown assays with MOB4 to identify the minimal binding region (e.g., MST4 amino acids 316-335) and critical phospho-acceptor residues (e.g., Thr-327/328) [6].
Biophysical Analysis of Binding Affinity: Bio-layer Interferometry (BLI) provides quantitative data on binding kinetics and affinity. This technique determined the dissociation constant (Kd) for the MST4-MOB4 interaction to be 1.67 µM, indicating a direct and stable complex formation [6].
Structural Determination: Although not detailed in the provided methodologies, the overall structural resemblance of the MST4-MOB4 complex to the MST1-MOB1 complex is confirmed through techniques like X-ray crystallography, as referenced by "structural studies" and "overall structure" [6].
The Scientist's Toolkit: Key Research Reagents
Table 3: Essential Reagents for MOB-Kinase Interaction Studies
| Reagent / Tool | Specific Example | Function in Experimental Protocol |
|---|---|---|
| Expression Vectors | SFB triple-tag (S-protein, FLAG, SBP) [17], MBP-fusion vectors [6] | For tandem affinity purification (TAP) and pulldown assays. |
| Cell Lines | HEK293FT, HEK293T, HEK293A, PANC-1 [6] [17] | Standard models for protein overexpression, interaction studies, and cancer-related phenotypic assays. |
| Phosphatase | λ Protein Phosphatase (λPP) [6] | To dephosphorylate proteins and test phosphorylation-dependence of interactions. |
| Kinase Mutants | Kinase-inactive (e.g., MST4-K53R) [6] | To determine the role of kinase activity and autophosphorylation in complex assembly. |
| Truncation Mutants | MST4 linker region mutants (e.g., Δ316-335) [6] | To map the minimal binding region and critical phospho-sites on the kinase. |
| Biophysical Instrument | Bio-layer Interferometer (BLI) [6] | To quantify binding affinity (Kd) and kinetics of protein-protein interactions. |
| Mass Spectrometry | TAP-MS, iTRAQ, PRM [18] [17] [19] | For unbiased identification of interaction partners and quantitative phosphoproteomics. |
Biological Implications and Pathophysiological Context
The functional divergence between MOB1 and MOB4 has direct consequences for cellular homeostasis and disease.
- Oncogenic Signaling: In pancreatic cancer, the MST4-MOB4 complex is upregulated and promotes cell growth and migration. In contrast, MST1 expression is often down-regulated. The MST4-MOB4 complex drives oncogenesis by disrupting the tumor-suppressive MST1-MOB1 complex, leading to YAP activation [6].
- Neuronal Development and Function: MOB4 is highly expressed in the central nervous system and is critical for neurodevelopment. It localizes to dendritic spines and is involved in synapse formation, microtubule organization, and axonal transport. Knockdown of MOB4 in zebrafish leads to severe neurologic defects, including a reduced hindbrain and eye size due to impaired cell division [7].
- The STRIPAK Connection: MOB4's role as a core component of the STRIPAK complex positions it as a key regulator of multiple signaling pathways. STRIPAK, through MOB4 and other subunits, directs the phosphatase PP2A toward specific targets like MAP4K4, thereby influencing Hippo signaling and contributing to oncogenic transformation [16].
This comparative analysis underscores that while MOB1 and MOB4 share a conserved structural fold, they have evolved distinct phospho-recognition interfaces that integrate them into opposing signaling circuits. MOB1 is a dedicated activator of the Hippo pathway's tumor-suppressive kinase cascade, whereas MOB4, via its integration into the STRIPAK complex and its ability to form an alternative complex with MST4, functions as a potent antagonist of Hippo signaling. The detailed molecular understanding of their binding interfaces and the functional consequences of their interactions provide a foundation for future research and the potential development of therapeutic strategies targeting these nodes in diseases like cancer and neurological disorders.
The Hippo signaling pathway is a conserved master regulator of tissue growth, organ size, and cellular homeostasis, with its dysregulation being a hallmark of cancer [4]. At the core of this pathway are kinase cascades where Mammalian STE20-like protein kinases (MSTs) and their adaptors, MOB proteins, play pivotal roles. The canonical MST1-MOB1 complex is a well-established tumor suppressor that potently inhibits the oncogenic transcriptional coactivator YAP (Yes-associated protein) [6] [4]. In contrast, emerging research reveals that the MST4-MOB4 complex, while structurally similar, has undergone functional divergence, acting as a non-canonical regulator with oncogenic potential [6] [14]. This divergence is critically framed within the broader context of Hippo signaling regulation, where MOB1 functions as a core pathway component and MOB4 acts as a key element of the STRIPAK complex, a known negative regulator of Hippo signaling [14] [7]. This guide provides a structured comparison of these functionally opposed complexes, detailing their mechanisms, experimental evidence, and implications for therapeutic targeting.
Biological Functions and Disease Roles
The MST1-MOB1 and MST4-MOB4 complexes exert opposing effects on cell proliferation, migration, and tumorigenesis, fundamentally influencing cancer progression.
Table 1: Comparative Biological Functions of MOB1 and MOB4 Complexes
| Feature | MST1-MOB1 Complex | MST4-MOB4 Complex |
|---|---|---|
| Primary Role | Tumor Suppressor [6] | Oncogenic Promoter [6] |
| Function in Hippo Pathway | Canonical activator of LATS1/2, leading to YAP phosphorylation/inactivation [4] | Non-canonical disruptor of MST1-MOB1, leading to YAP activation [6] |
| Effect on Cell Proliferation | Suppresses growth [20] [21] | Promotes growth [6] [22] |
| Effect on Cell Migration/Invasion | Inhibits migration [6] | Enhances migration and invasion [6] [22] |
| Role in Tumorigenesis | Restricts tumor development; loss promotes cancer [20] [21] | Drives tumor progression; overexpression is oncogenic [6] [22] |
| Expression in Cancer | Frequently down-regulated (e.g., pancreatic cancer) [6] | Frequently up-regulated (e.g., pancreatic, breast cancer) [6] [22] |
| Association with Patient Survival | Favorable prognosis [6] | Poor overall survival [22] |
| Key Regulatory Mechanism | Phosphorylation-triggered activation of LATS1/2 [4] [23] | Disruption of MST1-MOB1 complex; activation of alternative pathways (e.g., AKT) [6] [22] |
The MST4-MOB4 complex promotes oncogenesis through multiple mechanisms. In pancreatic cancer (PANC-1 cells), this complex synergistically enhances cell growth and migration [6]. Similarly, in breast cancer, MST4 overexpression accelerates cell growth, migration, and invasion, while its knockdown attenuates these properties [22]. MST4 also promotes Epithelial-Mesenchymal Transition (EMT), a key step in metastasis, by activating the AKT signaling pathway, resulting in decreased E-cadherin and increased N-cadherin, Snail, and Slug levels [22]. Furthermore, due to divergent evolution of key interface residues, MST4 and MOB4 disrupt the tumor-suppressive MST1-MOB1 complex through alternative pairing, thereby increasing YAP activity and driving oncogenic transformation [6].
Structural and Mechanistic Insights
Structural Conservation and Functional Divergence
Despite their opposing biological functions, the overall structures of the MST1-MOB1 and MST4-MOB4 complexes are remarkably similar [6] [24]. Both interactions are phosphorylation-dependent; MST kinases autophosphorylate at specific threonine residues in their linker regions, creating docking motifs for their respective MOB partners [6] [4].
The critical divergence lies in the functional outcome of these interactions. The MST1-MOB1 complex assembly is a key step in Hippo pathway activation. MOB1 exists in an autoinhibited state, where its N-terminal "Switch helix" blocks the LATS1-binding surface [23]. Phosphorylation of MOB1 by MST1 at Thr12 and Thr35 relieves this autoinhibition, inducing a conformational change that opens the LATS1-binding site [4] [23]. The activated phospho-MOB1 then binds to and facilitates the full activation of LATS1/2 kinases, which in turn phosphorylate and inhibit the oncoprotein YAP [4].
Conversely, the MST4-MOB4 complex disrupts this tumor-suppressive cascade. The formation of the MST4-MOB4 complex competitively interferes with the assembly of the MST1-MOB1 complex [6]. This disruption prevents the proper activation of the LATS1/2 kinases, leading to the accumulation of unphosphorylated, active YAP in the nucleus, where it drives the expression of pro-proliferative and anti-apoptotic genes [6]. Additionally, MOB4, as a core component of the STRIPAK complex, contributes to the dephosphorylation and inactivation of MST1/2 kinases, further antagonizing the canonical Hippo pathway [14] [7].
The following diagram illustrates the opposing signaling pathways governed by these complexes.
Key Experimental Evidence and Data
The functional divergence between these complexes is supported by a wealth of experimental data from biochemical, cellular, and clinical studies.
Table 2: Summary of Key Experimental Findings
| Complex | Experimental Model | Key Findings | Reference |
|---|---|---|---|
| MST1-MOB1 | Drosophila genetic studies | Loss of mats (MOB1) results in tissue overgrowth; acts downstream of hpo (MST1/2). | [20] |
| In vitro reconstitution & crystallography | Phospho-MST2 binds MOB1, relieving autoinhibition; phospho-MOB1 binds LATS1, enabling its activation. | [4] [23] | |
| MST4-MOB4 | Pancreatic cancer cells (PANC-1) | Complex promotes cell growth/migration; expression elevated in PC and negatively correlates with patient survival. | [6] |
| Breast cancer cells (MDA-MB-231, BT474) | MST4 overexpression promotes growth, migration, invasion, and EMT via AKT activation. | [22] | |
| X-ray crystallography (PDB: 5YF4) | Structure of MST4-MOB4 complex reveals basis for competitive disruption of MST1-MOB1. | [24] |
Detailed Experimental Protocols
To equip researchers with methodologies for investigating these complexes, this section outlines standard protocols derived from the cited literature.
Co-Immunoprecipitation (Co-IP) to Detect Complex Formation
This protocol is used to validate physical interactions between MST and MOB proteins in cells [6].
- Transfection: Culture HEK293FT or relevant cancer cells (e.g., PANC-1, MDA-MB-231) in appropriate medium. Transfect with plasmids expressing tagged versions of the proteins of interest (e.g., FLAG-MST4 and HA-MOB4) using a transfection reagent like Lipofectamine 2000.
- Cell Lysis: 24-48 hours post-transfection, lyse cells in a non-denaturing lysis buffer (e.g., RIPA buffer) supplemented with protease and phosphatase inhibitors.
- Immunoprecipitation: Incubate the cell lysate with an antibody specific to the tag of one protein (e.g., anti-FLAG M2 affinity gel) for several hours or overnight at 4°C with gentle rotation.
- Washing: Pellet the beads and wash extensively with cold lysis buffer to remove non-specifically bound proteins.
- Elution and Analysis: Elute the bound proteins by boiling in SDS-PAGE loading buffer. Analyze the eluates and input controls by Western blotting using antibodies against the tags of both binding partners (e.g., anti-HA to detect co-precipitated MOB4).
In Vitro Pulldown Assay with Purified Proteins
This method confirms a direct, phosphorylation-dependent interaction, excluding indirect cellular factors [6].
- Protein Purification: Express and purify recombinant proteins (e.g., MBP-tagged MST4 and His-tagged MOB4) from E. coli.
- Phosphorylation/Dephosphorylation Treatment:
- For phosphorylation: Incubate MBP-MST4 with ATP and MgCl₂.
- For dephosphorylation: Treat MBP-MST4 with λ protein phosphatase (λPP).
- Pulldown: Incubate the treated MBP-MST4 (or MBP control) with purified MOB4 in binding buffer. Add amylose resin to capture MBP-fused proteins.
- Washing and Elution: Wash the resin thoroughly to remove unbound MOB4. Elute the bound proteins with buffer containing maltose.
- Detection: Analyze the eluates by SDS-PAGE and Coomassie staining or Western blotting to detect the presence of MOB4, indicating a direct interaction.
Functional Cell-Based Assays
These assays quantify the biological impact of modulating MST-MOB complexes [6] [22].
- Cell Proliferation Assay:
- Generate stable cell lines with MST4/MOB4 overexpression or knockdown using lentiviral transduction.
- Seed cells in plates and count them every 24 hours for several days using a hemocytometer or automated cell counter. Plot cell number over time to assess growth rates.
- Migration and Invasion Assay (Boyden Chamber):
- Use a 24-well Transwell system with a porous (8 μm) membrane.
- For migration, seed serum-starved cells in the top chamber with a low-serum medium. Place medium with a serum gradient (chemoattractant) in the bottom chamber.
- For invasion, pre-coat the membrane with Matrigel matrix to simulate the extracellular matrix.
- After incubation (e.g., 8-24 hours), fix the cells that have migrated/invaded to the lower surface of the membrane, stain with crystal violet, and count under a microscope.
The Scientist's Toolkit: Key Research Reagents
Table 3: Essential Reagents for Studying MOB Complexes
| Reagent | Function/Application | Example |
|---|---|---|
| Expression Plasmids | Overexpression or mutant analysis of MST/MOB genes. | FLAG-MST4, HA-MOB4, MST4 kinase-dead (K53R, T178A) mutants [6] [22]. |
| Knockdown Tools | Loss-of-function studies to assess necessity. | shRNA lentiviral particles targeting MST4 or MOB4 [22]. |
| Phospho-Specific Antibodies | Detecting activation-specific phosphorylation events. | Anti-pMOB1 (Thr12/Thr35) [4]; anti-pYAP [4]. |
| Structural Biology Tools | Determining atomic-level interaction mechanisms. | Purified MST4-MOB4 complex for crystallization (PDB: 5YF4) [24]. |
| Cell Line Models | Studying context-specific functions in disease. | Pancreatic cancer: PANC-1 [6]. Breast cancer: MDA-MB-231, BT474 [22]. |
The functional divergence between the tumor-suppressive MST1-MOB1 complex and the oncogenic MST4-MOB4 complex highlights the intricate regulatory landscape of the Hippo pathway. While the former serves as the canonical engine for growth inhibition, the latter represents a potent non-canonical bypass that promotes tumorigenesis through direct disruption and alternative signaling. This comparison underscores the critical importance of understanding specific MST-MOB pairings in cancer biology. Targeting the oncogenic MST4-MOB4 interface or restoring the tumor-suppressive MST1-MOB1 function presents a promising, albeit challenging, therapeutic strategy for cancers driven by YAP/TAZ activation. Future research should focus on elucidating the precise regulatory signals that dictate the balance between these complexes and developing selective inhibitors against the MST4-MOB4 axis.
The Mps one binder (MOB) family represents a group of highly conserved, non-catalytic adaptor proteins that serve as critical regulators of kinase signaling pathways. Within this family, MOB1 and MOB4 have emerged as key players with distinct, often opposing, biological functions despite sharing structural similarities. MOB1 is well-established as a core component of the canonical Hippo pathway, acting as a tumor suppressor by restricting organ growth and cell proliferation. In contrast, MOB4, initially identified as Phocein, functions primarily within the Striatin-interacting phosphatase and kinase (STRIPAK) complex and demonstrates oncogenic properties in certain contexts [8] [7]. This comparison guide objectively analyzes the differential roles of MOB1 and MOB4 in Hippo signaling, drawing on structural, functional, and mechanistic data to provide researchers with a clear framework for understanding their distinct cellular functions.
Table 1: Fundamental Classification of MOB1 and MOB4
| Feature | MOB1 | MOB4 |
|---|---|---|
| Primary Complex | Hippo signaling core | STRIPAK complex |
| Evolutionary Class | Class I MOB | Class IV MOB |
| Key Alias Names | MOBKL1A/B, Mats | Phocein, MOBKL3 |
| Conservation with Drosophila | dMOB1 (85% identity) | dMOB4 (80% identity) |
| Domain Structure | Conserved Mob/Phocein domain | Conserved Mob/Phocein domain with divergent N-terminal |
Structural and Molecular Characteristics
Shared Structural Fold with Distinct Interfaces
Both MOB1 and MOB4 proteins adopt the conserved globular Mob/Phocein domain fold, characterized by a four-helix bundle at its core with three short α-helices at the N-terminal extension [8] [7]. This shared structural foundation, however, belies significant functional divergence. While MOB1 maintains the capacity for stable binding to Nuclear Dbf2-Related (NDR) kinases including LATS1/2, MOB4 lacks this ability and has evolved distinct protein interaction interfaces [8]. The human MOB4 gene generates multiple alternatively spliced transcriptional variants, producing three predicted protein isoforms that may contribute to its functional versatility [7].
Differential Binding Specificities
The critical functional distinction arises from their differential binding specificities for STE20-like kinases. MOB1 forms a stable complex with MST1/2 kinases in a phosphorylation-dependent manner, serving as an essential adaptor that facilitates LATS1/2 activation [6]. Conversely, MOB4 specifically interacts with MST4 kinase, also in a phosphorylation-dependent fashion, with a dissociation constant (Kd) of approximately 1.67 μm [6]. This interaction depends on MST4 autophosphorylation at Thr-327/328 within its linker region, which creates a binding interface for MOB4 [6]. Despite structural similarities between the MST1-MOB1 and MST4-MOB4 complexes, they exert opposing biological effects, highlighting their functional divergence.
Signaling Mechanisms and Pathway Regulation
MOB1 in Canonical Hippo Signaling
MOB1 functions as a critical signal transducer in the canonical Hippo tumor suppressor pathway. Upon activation, MST1/2 kinases phosphorylate MOB1, which subsequently interacts with and fully activates LATS1/2 kinases [6]. The activated LATS1/2 complex then phosphorylates the transcriptional coactivators YAP/TAZ, leading to their cytoplasmic retention and proteolytic degradation [6]. This signaling cascade effectively suppresses the expression of proliferative and anti-apoptotic genes, positioning the MST1-MOB1 complex as a fundamental growth-restricting module in cells.
MOB4 in STRIPAK-Mediated Pathway Regulation
MOB4 operates primarily as a core component of the STRIPAK complex, which includes protein phosphatase 2A (PP2A), striatins, and various other regulatory subunits [7] [11]. Within this complex, MOB4 participates in the negative regulation of Hippo signaling through dephosphorylation of MST kinases. The STRIPAK complex serves as a supramolecular assembly that brings PP2A phosphatase into proximity with its kinase substrates, enabling precise control of phosphorylation states [11]. Recent structural studies have revealed that STRN3 acts as an organizing center within STRIPAK, with PP2Aα/c-bound STRN3 directly contacting Hippo kinases and controlling their loading via two "arms" - one being STRIP1 and the other SIKE1-SLMAP [11].
Diagram 1: MOB1 and MOB4 in opposing signaling pathways. MOB1 activates Hippo tumor suppressor signaling while MOB4-containing STRIPAK complex inhibits it.
Antagonistic Functional Relationship
A significant mechanistic insight reveals that MST4-MOB4 complex directly disrupts MST1-MOB1 complex assembly through alternative pairing, thereby increasing YAP activity [6]. This functional antagonism arises from evolutionary divergence of key interface residues that enable competitive binding interactions. The MST4-MOB4 complex promotes YAP-driven transcription by sequestering components required for canonical Hippo pathway activation, representing a non-canonical regulatory mechanism within the broader Hippo signaling network [6] [25].
Table 2: Functional Outcomes in Cellular Contexts
| Cellular Process | MOB1 Role | MOB4 Role |
|---|---|---|
| Pancreatic Cancer | Tumor suppressor: inhibits growth and migration | Oncogenic: promotes growth and migration |
| Expression in Pancreatic Cancer | Down-regulated | Up-regulated, correlated with poor survival |
| Neurogenesis | Limited direct role | Essential: regulates neuronal branching, microtubule organization |
| Neurite Development | Not reported | Critical: knockout causes hyperbranching and defective connections |
| Schwann Cell Development | Not well characterized | Required for radial sorting and YAP/TAZ regulation |
Experimental Approaches and Key Findings
Protein Interaction Studies
Co-immunoprecipitation assays in HEK293FT cells have demonstrated that FLAG-tagged MST4 readily pulls down HA-tagged MOB4, confirming their physical interaction [6]. Endogenous co-IP experiments further validated this association under physiological conditions. For direct binding assessment, recombinant protein pulldown assays using purified MST4 and MOB4 proteins expressed in E. coli confirmed their specific interaction, with MBP-tagged MST4 successfully pulling down MOB4 but not MBP control [6]. Bio-layer interferometry quantified this interaction, revealing a Kd of 1.67 μm [6].
To determine the structural basis of MST4-MOB4 interaction, researchers employed crystallographic approaches that demonstrated the overall structure of the MST4-MOB4 complex resembles that of the MST1-MOB1 complex, despite their functional differences [6] [25]. Phosphorylation-dependency experiments using λ protein phosphatase treatment showed that dephosphorylated MST4 loses MOB4 binding capacity, which can be restored through auto-phosphorylation with ATP [6]. Truncation mutagenesis identified amino acids 316-335 of the MST4 linker region as the minimal binding region for MOB4 [6].
Functional Characterization in Disease Models
In pancreatic cancer models, overexpression and knockdown studies revealed opposing functions: while MST1-MOB1 complex suppressed oncogenic phenotypes, MST4-MOB4 complex promoted PANC-1 cell growth and migration [6]. Immunohistochemical analysis of patient samples showed elevated MST4 and MOB4 expression in pancreatic tumors, positively correlating with each other while MST1 expression was down-regulated [6]. Survival analyses established clinical relevance, connecting MST4-MOB4 upregulation with negative patient outcomes [6].
In neurological contexts, genetic knockout models in Drosophila demonstrated that Mob4 is essential for viability, with null mutants not surviving past larval stages [7]. Tissue-specific knockdown revealed Mob4's critical role in neuronal development, with deficient neurons showing abnormal branching patterns, disrupted microtubule organization, and defective synaptic development [7]. Morpholino-mediated knockdown in zebrafish embryos caused severe neurologic defects, including loss of midbrain-hindbrain boundary and reduced eye size, establishing its evolutionary conserved role in neurogenesis [7].
Research Reagent Solutions
Table 3: Essential Research Tools for MOB1/MOB4 Investigation
| Reagent Category | Specific Examples | Research Application |
|---|---|---|
| Expression Plasmids | FLAG-MST4, HA-MOB4, MBP-tagged truncation mutants | Protein interaction studies, pulldown assays |
| Cell Lines | HEK293FT, PANC-1 | Interaction validation, functional assays in cancer models |
| Antibodies | Anti-MST4, Anti-MOB4, Phospho-specific MOB1 | Endogenous co-IP, localization, activation status |
| Recombinant Proteins | Purified MST4, MOB4, λ protein phosphatase | Direct binding assays, phosphorylation dependency |
| Kinase Assay Components | ATP, MgCl₂, kinase-inactive mutants (MST4-K53R) | Phosphorylation studies, functional characterization |
| Animal Models | Drosophila Mob4 mutants, Zebrafish morphants | Neurological function, developmental roles |
The comparative analysis of MOB1 and MOB4 reveals two structurally related adaptor proteins with fundamentally opposing biological functions within cellular signaling networks. While MOB1 acts as a cornerstone of the tumor-suppressive Hippo pathway, MOB4 functions within the STRIPAK complex to antagonize Hippo signaling and promote growth and migration in specific contexts. Their differential expression patterns in diseases such as pancreatic cancer, coupled with their specialized roles in processes like neurogenesis, highlight the complexity of cellular regulation and offer potential avenues for therapeutic intervention. Future research should focus on elucidating the precise structural determinants of their binding specificities and exploring the therapeutic potential of modulating these interactions in cancer and neurological disorders.
Techniques for Mapping MOB Interactions and Functional Outcomes in Hippo Signaling
Proximity-Dependent Biotin Identification (BioID) to Elucidate MOB Proximity Interactomes
Proximity-dependent biotin identification (BioID) has emerged as a transformative methodology for mapping protein-protein interactions (PPIs) in living cells, enabling researchers to capture transient, weak, and spatially constrained interactions that evade traditional biochemical methods. This technique has proven particularly valuable for elucidating the interactomes of adaptor proteins, including the highly conserved Mps one binder (MOB) family, which play critical roles in key signaling pathways such as Hippo and STRIPAK. This guide provides a comprehensive comparison of BioID applications for mapping MOB protein proximity networks, with a specialized focus on differentiating the interactomes of canonical MOB1 and non-canonical MOB4. We present structured experimental data, detailed methodologies, and essential resource information to equip researchers with the tools necessary to investigate these crucial regulatory proteins in physiological and pathological contexts.
BioID utilizes a promiscuous mutant of the Escherichia coli biotin ligase (BirA*, R118G) fused to a protein-of-interest. This fusion protein catalyzes the covalent biotinylation of proximate proteins (within ~10 nm) in the presence of exogenous biotin, generating a history of protein associations over time [26]. The biotinylated proteins can subsequently be purified under denaturing conditions using streptavidin affinity capture and identified via mass spectrometry (MS) [26] [27]. This approach offers several distinct advantages for mapping MOB protein interactions: (1) ability to capture weak and transient interactions, (2) applicability to insoluble cellular compartments and membrane-associated proteins, (3) temporal control through biotin supplementation, and (4) preservation of cellular context during labeling [26] [28].
The labeling radius of BioID has been experimentally validated at approximately 10 nm, representing one of the strictest labeling distances among proximity-dependent labeling methods [26] [29]. Biotinylated proteins identified through BioID typically fall into three categories: direct interactors (both transient and stable), indirect interactors, and proximal proteins that do not physically interact but reside within the labeling vicinity [26]. This technology has been successfully applied across diverse systems, including mammalian cells, plants, and mice, demonstrating its broad utility [26] [28].
Comparative Analysis of MOB1 and MOB4 Proximity Interactomes
MOB proteins function as critical adaptors in cellular signaling networks. The Class I MOB1 proteins are well-established components of the Hippo pathway, whereas Class IV MOB4 acts as a core constituent of the Striatin-interacting phosphatase and kinase (STRIPAK) complex, which negatively regulates Hippo signaling [8] [7]. Systematic BioID screening of all seven human MOB proteins in HeLa and HEK293 cell lines has revealed distinct interaction profiles for MOB1 and MOB4, illuminating their specialized cellular functions [30].
Table 1: MOB1 vs. MOB4 Proximity Interactomes Identified by BioID
| Feature | MOB1 (Class I) | MOB4 (Class IV) |
|---|---|---|
| Primary Signaling Pathway | Core Hippo pathway [8] | STRIPAK complex [7] |
| Key Interactors | LATS1/2, STK3/4 (MST1/2), SAV1, PP6 complex [30] | STRNs, MST4, PP2A phosphatases, STK38/STK38L (in specific contexts) [6] [12] |
| Validated Functional Role | Tumor suppressor; activates LATS1/2 to inhibit YAP/TAZ [8] | Regulates cytoskeletal dynamics, neuronal development; can promote oncogenic signaling in specific contexts [12] [7] |
| Subcellular Localization | Cytoplasmic, associated with plasma membrane [8] | Somatodendritic in neurons, Golgi apparatus, associated with cytoskeleton [6] [7] |
| Percentage of Novel Interactions | ~27% previously unreported in BioGrid [30] | >70% previously unreported in BioGrid [30] |
Table 2: Functional Consequences of MOB Protein Interactions
| Cellular Process | MOB1-Dependent Regulation | MOB4-Dependent Regulation |
|---|---|---|
| Cell Proliferation | Suppresses proliferation via YAP/TAZ inactivation [8] | Promotes proliferation through STRIPAK-mediated Hippo inhibition [7] |
| Neuronal Development | Limited direct role | Essential for dendritic spine formation, microtubule organization, and synaptic development [7] |
| Tumorigenesis | Tumor suppressor; frequently downregulated in cancer [8] | Often overexpressed in tumors; linked to poor clinical outcomes [7] |
| Complex Disruption | MST1-MOB1 disrupted by alternative MST4-MOB4 pairing [6] | MST4-MOB4 competes with MST1-MOB1 complex formation [6] |
Experimental Design and Protocol for MOB BioID Screening
Vector Construction and Cell Line Generation
For systematic MOB interactome mapping, researchers have successfully employed N-terminal tagging strategies with BirA-FLAG-MOB fusions cloned into tetracycline-inducible expression vectors [30]. This configuration positions the BirA enzyme optimally for proximity labeling while maintaining the native structure and function of MOB proteins. The inducible expression system is crucial for controlling the timing and level of fusion protein expression, minimizing potential artifacts from constitutive overexpression.
Essential Controls: The experimental design must include appropriate negative controls, typically BirA-FLAG or BirA-FLAG-EGFP, to account for background biotinylation and non-specific streptavidin binding [30]. These controls enable statistical discrimination of true proximal interactors from background proteins during mass spectrometry data analysis.
Stable Cell Line Generation: Both HEK293 and HeLa Flp-In T-REx cell lines have been successfully utilized for MOB BioID studies [30]. Stable integration ensures consistent expression levels and enables reproducible experiments across biological replicates, which is essential for robust statistical analysis.
BioID Labeling and Biotinylation Optimization
The BioID procedure requires careful optimization of biotin concentration and labeling duration to maximize signal-to-noise ratio while maintaining cellular viability:
Biotin Supplementation: Cells expressing BirA*-MOB fusions are treated with 50 μM biotin for 15-18 hours to achieve optimal labeling [26]. This duration allows sufficient accumulation of biotinylated proteins while maintaining cell viability.
Biotin Stock Preparation: Prepare 1 mM biotin stock solution in serum-free cell culture medium, filter-sterilize through 0.22 μM filters, and store at 4°C for up to 8 weeks [26].
Temporal Considerations: The optimal biotin incubation period represents a balance between sufficient labeling and potential toxicity. For MOB proteins, 15-18 hour labeling periods have proven effective for capturing both stable and transient interactions [30].
Streptavidin Affinity Purification and Protein Identification
Following biotinylation, cells are lysed under denaturing conditions (e.g., using RIPA buffer with SDS) to disrupt all non-covalent interactions while preserving the covalent biotin tags [26] [30]. The biotinylated proteins are then captured using streptavidin-conjugated beads and subjected to stringent washing to remove non-specifically bound proteins.
Critical Steps:
- Use high-capacity streptavidin-agarose beads for efficient capture
- Include multiple wash steps with SDS-containing buffers to reduce background
- Perform on-bead trypsin digestion for mass spectrometry analysis
- Analyze peptides by LC-MS/MS using high-resolution instruments
Data Analysis: Process MS data using standard proteomics pipelines and apply statistical frameworks such as Significance Analysis of INTeractome (SAINT) to identify high-confidence interactors with controlled false discovery rates [30] [28].
Signaling Pathway Diagrams
Diagram 1: MOB1-Hippo and MOB4-STRIPAK Signaling Networks. The canonical MOB1-dependent Hippo pathway (red) suppresses YAP/TAZ activity and cell proliferation, while the MOB4-STRIPAK complex (blue) promotes YAP/TAZ activation. These pathways exhibit mutual antagonism through competitive interactions between MST-MOB complexes [6] [8] [7].
Diagram 2: BioID Experimental Workflow for MOB Interactome Mapping. The complete procedure from vector construction to data analysis, highlighting critical control elements and statistical validation steps essential for generating high-confidence proximity interactomes [26] [30] [27].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for MOB BioID Studies
| Reagent Category | Specific Examples | Function/Application | Technical Notes |
|---|---|---|---|
| Expression Vectors | BirA*-FLAG (N-terminal tagging); Tetracycline-inducible systems [30] | Inducible expression of BirA*-MOB fusions | N-terminal tagging preserves MOB protein function; inducible systems prevent artifacts |
| Cell Lines | HEK293 Flp-In T-REx; HeLa Flp-In T-REx [30] | Stable, inducible expression platforms | Ensure proper subcellular localization of BirA*-MOB fusions |
| Biotin & Supplements | 1 mM biotin stock (50 μM working concentration) [26] | Activates BirA*-mediated proximity labeling | Filter sterilize; avoid extended storage; optimize concentration for specific cell types |
| Detection Antibodies | α-FLAG, α-BioID (BID-CP-100), α-BioID2 (BID2-CP-100) [26] | Fusion protein detection and validation | Verify expression levels and biotinylation efficiency |
| Streptavidin Reagents | Streptavidin-conjugated beads, HRP, Alexa Fluors [26] [30] | Capture, detection, and visualization of biotinylated proteins | High-capacity beads improve yield; multiple fluorophores enable multiplexing |
| Mass Spectrometry | High-resolution LC-MS/MS systems [30] [28] | Identification of biotinylated peptides | Tandem MS enables protein identification and modification mapping |
| Analysis Software | SAINT, MaxQuant, MoSS algorithm [30] [28] | Statistical analysis of interaction data | Specificity scoring differentiates true interactors from background |
Discussion and Research Applications
The application of BioID to MOB protein interactome mapping has revealed unprecedented insights into the functional specialization of these adaptor proteins. The comprehensive BioID screening of all seven human MOB proteins identified over 200 interactions, with at least 70% representing previously unreported associations in BioGrid [30]. This expansion of the MOB interaction landscape underscores the power of proximity labeling technologies for discovering novel biology, particularly for poorly characterized proteins like MOB3C, which was found to associate with 7 of 10 protein subunits of the RNase P complex [30] [29].
The comparative analysis of MOB1 and MOB4 interactomes highlights their opposing functions in cellular signaling. While MOB1 serves as a tumor suppressor within the Hippo pathway, MOB4 can promote oncogenic signaling through its role in the STRIPAK complex, particularly in cancers such as pancreatic cancer where the MST4-MOB4 complex is upregulated and associated with poor prognosis [6] [25]. This functional divergence occurs despite structural conservation between MOB1 and MOB4, emphasizing how subtle differences in interaction preferences can translate to significant biological outcomes [7].
The integration of BioID-derived interaction data with functional studies provides a powerful approach for elucidating molecular mechanisms. For example, MOB4's role in neuronal development, particularly in dendritic spine formation and microtubule organization, has been clarified through combination of proximity labeling with genetic approaches in Drosophila and zebrafish [7]. Similarly, the competitive relationship between MST1-MOB1 and MST4-MOB4 complexes in regulating YAP/TAZ activity demonstrates how proximity interactomes can reveal previously unappreciated regulatory mechanisms [6].
Future applications of BioID in MOB research could include temporal mapping of interactome dynamics during cellular processes such as differentiation, cell cycle progression, or stress response. Additionally, the development of more sensitive biotin ligases (e.g., BioID2, TurboID) may enable faster labeling and detection of more transient interactions, further refining our understanding of MOB protein networks in health and disease [26].
The specificity of pairing between Mammalian Ste20-like (MST) kinases and Mps One Binder (MOB) adaptor proteins represents a critical regulatory node in cellular signaling, governing processes from tissue growth to tumor suppression. Within the canonical Hippo pathway, the MST1/2-MOB1 complex acts as a well-characterized tumor suppressor, activating LATS kinases to inhibit oncogenic YAP signaling [8] [31]. In contrast, emerging research reveals that the MST4-MOB4 complex, a component of the Striatin Interacting Phosphatase and Kinase (STRIPAK) complex, can disrupt this tumor-suppressive function and promote oncogenic activity in cancers such as pancreatic cancer [6] [25]. This comparison guide examines how co-immunoprecipitation (Co-IP) and pull-down assays serve as essential tools for delineating the specificity, affinity, and functional outcomes of these distinct MST-MOB pairings, providing researchers with methodological frameworks for investigating these biologically significant interactions.
Biological Background: Competitive MST-MOB Complexes
Canonical Hippo Signaling: The MST1-MOB1 Complex
The MST1-MOB1 complex forms a core signaling module within the conserved Hippo tumor suppressor pathway. This interaction is phosphorylation-dependent, with MST1 phosphorylating MOB1 to create a high-affinity binding surface that subsequently recruits and activates LATS1/2 kinases [32] [31]. Activated LATS kinases then phosphorylate the transcriptional co-activators YAP/TAZ, leading to their cytoplasmic retention and degradation. Structurally, MOB1 contains a conserved phosphopeptide-binding infrastructure that specifically recognizes phosphorylated motifs in MST1/2 [32]. The functional significance of this interaction is profound: MOB1 binding to LATS1/2 is essential for tumor suppression, tissue growth control, and proper development, while stable MOB1 interaction with MST1/2 itself appears dispensable for these functions [31].
Non-Canonical Signaling: The MST4-MOB4 Complex in STRIPAK
The MST4-MOB4 complex represents a non-canonical pairing with opposing biological functions to the MST1-MOB1 complex. MST4 directly interacts with MOB4 in a phosphorylation-dependent manner at Thr-327/328 within its linker region [6]. Unlike MOB1, MOB4 is a core component of the STRIPAK complex, a multi-subunit regulator of cellular phosphorylation states. Structurally, while the overall architecture of the MST4-MOB4 complex resembles that of MST1-MOB1, divergent evolution at key interface residues enables functional specialization [6]. Crucially, the MST4-MOB4 complex exhibits oncogenic properties in pancreatic cancer, promoting cell proliferation and migration rather than suppression [6] [25].
Competitive Disruption Between Complexes
The competitive relationship between these complexes represents a key regulatory mechanism. Research demonstrates that MST4 and MOB4 can disrupt assembly of the tumor-suppressive MST1-MOB1 complex through alternative pairing, thereby increasing YAP activity and driving oncogenic progression [6]. This competition occurs due to shared structural features combined with specific interface variations that enable cross-pairing while generating opposite functional outcomes in tumorigenesis.
Experimental Approaches: Co-IP and Pull-Down Assays
Co-Immunoprecipitation (Co-IP) for MST-MOB Interactions
Co-immunoprecipitation enables researchers to capture native protein complexes from cell lysates under conditions that preserve physiological interactions, making it ideal for studying MST-MOB partnerships in their cellular context.
Key Protocol for MST-MOB Co-IP
The fundamental Co-IP protocol involves several critical stages [33] [34]:
- Cell Lysis: Use freshly prepared non-denaturing lysis buffer (e.g., RIPA or NP-40) supplemented with protease and phosphatase inhibitors to preserve phosphorylation-dependent interactions. For Hippo pathway studies, maintain non-denaturing conditions to keep protein complexes intact [33].
- Antibody Incubation: Incubate cell lysate (typically 300μg-2mg total protein) with 2μg of target-specific antibody (anti-MST1, MST4, MOB1, or MOB4) for 1 hour at 4°C with rotation [34].
- Bead Capture: Add Protein A/G sepharose or magnetic beads (50% slurry) and incubate for an additional 1-3 hours at 4°C. Magnetic beads are preferred for minimizing mechanical disruption of complexes [35].
- Washing: Pellet beads by centrifugation and wash 3-4 times with ice-cold lysis buffer to remove non-specifically bound proteins. Remove supernatant carefully by pipetting rather than vacuum aspiration to prevent bead loss [33].
- Elution: Boil beads in reducing SDS-PAGE sample buffer for 5-10 minutes to dissociate immunoprecipitated complexes [34].
- Analysis: Subject eluates to SDS-PAGE followed by Western blotting with antibodies against putative interacting partners, or to mass spectrometry for interaction partner discovery [33].
Critical Controls for Interaction Specificity
Appropriate controls are essential for validating specific MST-MOB interactions [34]:
- Input Control: Reserve 1-10% of initial lysate to confirm presence of target proteins.
- Negative IgG Control: Use non-specific IgG from the same species to identify non-specific binding to beads or antibodies.
- Competition Control: Include phosphopeptide competitors to verify phosphorylation-dependent interactions, particularly important for MST-MOB1 binding [32].
- Genetic Controls: When available, use knockout cell lines or RNAi-mediated knockdown to confirm antibody specificity.
Pull-Down Assays for Direct Interaction Mapping
Pull-down assays employ recombinant affinity-tagged "bait" proteins to capture "prey" proteins from cell lysates or among purified components, providing complementary data to Co-IP by testing direct interactions without antibody mediation.
Key Protocol for MST-MOB Pull-Down
The standard pull-down approach includes [36]:
- Bait Preparation: Express and purify MST kinases (MST1, MST4) or MOB adaptors (MOB1, MOB4) as fusion proteins with affinity tags (GST, MBP, or His₆) from E. coli or mammalian expression systems.
- Immobilization: Incubate purified bait protein with appropriate affinity resin (glutathione-sepharose for GST, amylose resin for MBP, or nickel-NTA for His₆ tags) for 30-60 minutes at 4°C.
- Binding Reaction: Incubate immobilized bait with either purified prey protein or cell lysate containing potential interaction partners for 1-3 hours at 4°C with gentle shaking.
- Washing: Pellet resin by centrifugation and wash 3-4 times with binding buffer containing 150-300mM NaCl to remove weakly associated proteins.
- Elution: Competitively elute bound complexes using tag-specific eluents (glutathione for GST, imidazole for His₆, or maltose for MBP) or directly by boiling in SDS-PAGE buffer.
- Analysis: Detect bound interaction partners by Western blotting or mass spectrometry.
Phosphorylation-Dependency Assessment
For MST-MOB interactions, particularly MST1-MOB1 and MST4-MOB4, assessing phosphorylation dependence is crucial [32] [6]:
- Phosphatase Treatment: Treat bait proteins with λ protein phosphatase prior to pull-down to abolish phosphorylation-dependent interactions.
- Kinase-Inactive Mutants: Use kinase-dead MST variants (e.g., MST4-K53R) that cannot autophosphorylate, serving as negative controls.
- ATP Supplementation: Include ATP and MgCl₂ in binding reactions to support kinase activity and autophosphorylation.
Comparative Data: MST-MOB Interaction Profiles
Quantitative Binding Parameters
Table 1: Quantitative Binding Parameters of MST-MOB Complexes
| Interaction Pair | Dissociation Constant (Kd) | Phosphorylation Dependence | Structural Features | Functional Outcome |
|---|---|---|---|---|
| MST1-MOB1 | ~0.2-0.5 μM [31] | Strictly dependent on MST1-mediated MOB1 phosphorylation [32] | MOB1 phosphopeptide-binding domain recognizes phosphorylated MST1 linker region [32] | Tumor suppressive: Activates LATS1/2 to inhibit YAP [31] |
| MST4-MOB4 | ~1.67 μM [6] | Dependent on MST4 autophosphorylation at Thr327/328 [6] | Resembles MST1-MOB1 structure despite low sequence conservation [6] | Oncogenic: Promotes cell proliferation and migration [6] |
| MOB1-LATS1 | Not quantified | Enhanced by MOB1 phosphorylation at T12/T35 [31] | Asp63 of MOB1 specifically bonds with His646 of LATS1 [31] | Essential for tumor suppression and growth control [31] |
Functional Comparison in Disease Contexts
Table 2: Functional Properties of MST-MOB Complexes in Disease Models
| Parameter | MST1-MOB1 Complex | MST4-MOB4 Complex |
|---|---|---|
| Pathway Association | Core Hippo tumor suppressor pathway [8] [31] | STRIPAK complex regulatory module [6] [25] |
| Role in Pancreatic Cancer | Down-regulated; loss correlates with YAP activation [6] | Up-regulated; promotes growth and migration [6] |
| Interaction Specificity Determinants | MOB1 phospho-recognition domain; MST1 phosphorylation sites [32] | MST4 linker region (aa 316-335); phosphorylation at Thr327/328 [6] |
| Competitive Behavior | Disrupted by MST4-MOB4 alternative pairing [6] | Competes with MST1-MOB1 complex formation [6] |
| Therapeutic Implications | Potential activation strategy for tumor suppression [31] | Potential inhibition target for oncology [6] |
Technical Considerations for Assay Selection
Co-IP vs. Pull-Down: Advantages and Limitations
The choice between Co-IP and pull-down assays depends on specific research questions and available reagents. Co-IP excels at capturing physiological interactions from native cellular environments but depends heavily on antibody quality and specificity [34]. Pull-down assays provide more controlled conditions for mapping direct interactions and are indispensable when high-quality IP-validated antibodies are unavailable [36]. For comprehensive MST-MOB interaction analysis, both techniques should be employed as complementary approaches.
Optimization Strategies for MST-MOB Studies
Successful validation of MST-MOB pairing specificity requires careful optimization:
- Lysis Conditions: Use non-denaturing buffers (e.g., 20mM Tris pH 7.5, 150mM NaCl, 1% NP-40) with fresh protease and phosphatase inhibitors to preserve labile phosphorylation-dependent interactions [33] [34].
- Bead Selection: Protein A/G beads are standard for Co-IP, while glutathione-sepharose (GST-tag), nickel-NTA (His-tag), or streptavidin beads (biotin-tag) are common for pull-downs. Magnetic beads minimize mechanical disruption during washes [33] [35].
- Wash Stringency: Optimize wash buffer stringency (150-500mM NaCl) to balance signal-to-noise ratio while preserving specific interactions. For weak or transient interactions, use lower salt concentrations (150mM NaCl).
- Detection Sensitivity: For low-abundance endogenous complexes, consider sensitive detection methods such as enhanced chemiluminescence or proximity ligation assays.
Essential Research Reagents and Tools
Table 3: Key Reagent Solutions for MST-MOB Interaction Studies
| Reagent Category | Specific Examples | Application Notes |
|---|---|---|
| Antibodies for Co-IP | Anti-MST1, Anti-MST4, Anti-MOB1, Anti-MOB4 [33] | Must be validated for immunoprecipitation; check species reactivity |
| Affinity Beads | Protein A/G agarose, Magnetic Dynabeads [35] | Magnetic beads preferred for minimizing complex disruption |
| Lysis Buffers | Non-denaturing RIPA, NP-40-based buffers [33] [34] | Supplement with fresh phosphatase inhibitors for phosphorylation-dependent interactions |
| Expression Vectors | GST-MST1, GST-MST4, His-MOB1, His-MOB4 [6] [31] | Enable recombinant protein production for pull-down assays |
| Phosphatase Inhibitors | Sodium fluoride, β-glycerophosphate, Sodium orthovanadate [33] | Critical for preserving phosphorylation-dependent interactions |
| Protease Inhibitors | PMSF, Complete Mini tablets [33] | Prevent protein degradation during lysate preparation and IP |
Co-immunoprecipitation and pull-down assays provide complementary and powerful approaches for delineating the specificity of MST-MOB pairings in both Hippo and STRIPAK signaling contexts. The competitive relationship between tumor-suppressive MST1-MOB1 and oncogenic MST4-MOB4 complexes highlights the importance of rigorous interaction validation using the methodological frameworks outlined in this guide. As research continues to unravel the complexity of MST-MOB networks, these fundamental techniques will remain essential for understanding their roles in development, tissue homeostasis, and disease pathogenesis, ultimately informing therapeutic strategies targeting these critical regulatory interactions.
The Hippo signaling pathway is an evolutionarily conserved kinase cascade that plays a crucial role in controlling organ size, tissue homeostasis, and cell proliferation [2]. Central to this pathway is the regulation of the transcriptional coactivators YAP (Yes-associated protein) and TAZ (transcriptional coactivator with PDZ-binding motif), whose activity is predominantly controlled by the LATS1/2 (large tumor suppressor kinase 1/2) kinases [37] [2]. The activity of LATS1/2 is, in turn, regulated by distinct MOB (Monopolar spindle-one-binder) protein complexes, creating a critical junction in Hippo pathway signaling.
The canonical MOB1 complex functions as a direct activator of LATS1/2 kinases. When the Hippo pathway is activated, MST1/2 kinases phosphorylate and activate MOB1, which then binds to and facilitates LATS1/2 activation [2]. Activated LATS1/2 subsequently phosphorylates YAP/TAZ on multiple serine residues, leading to their cytoplasmic retention and proteasomal degradation [38].
In contrast, the MOB4 complex acts as a negative regulator of Hippo signaling through its role in the STRIPAK (Striatin-interacting phosphatase and kinase) complex [9] [7]. STRIPAK is a large multisubunit PP2A phosphatase complex that includes MOB4, STRN3 (Striatin-3), STRIP1/2, and the PP2A catalytic and scaffold subunits [9]. This complex dephosphorylates and inhibits MST1/2 kinases, thereby suppressing the kinase cascade that would otherwise lead to LATS1/2 activation and YAP/TAZ inhibition [7].
This comparison guide provides an objective evaluation of experimental methods for measuring LATS1/2 and YAP/TAZ phosphorylation, with a specific focus on the distinct regulatory functions of MOB1 versus MOB4 complexes in Hippo pathway research.
Key Signaling Nodes: LATS1/2 and YAP/TAZ Phosphorylation
LATS1/2 Kinase Activation and Substrate Phosphorylation
LATS1 and LATS2 are serine/threonine kinases belonging to the AGC kinase family that serve as the central signaling node where MOB complexes exert their regulatory influence [2]. Activation of LATS kinases requires phosphorylation at specific sites: LATS1 at S909 and LATS2 at S872 in their activation loops [39]. This phosphorylation can be catalyzed by upstream kinases including MST1/2 and MAP4Ks [2]. Once activated, LATS1/2 directly phosphorylates YAP and TAZ on multiple conserved residues.
The functional assessment of LATS kinase activity is typically measured through:
- Autophosphorylation status at activation loop sites (LATS1 pS909, LATS2 pS872)
- Transphosphorylation capability against substrate proteins, particularly YAP/TAZ
- Cellular localization and complex formation with MOB proteins
YAP/TAZ Phosphorylation and Functional Consequences
YAP and TAZ are the primary downstream effectors of the Hippo pathway and serve as key substrates for LATS1/2 kinases. LATS-mediated phosphorylation occurs on multiple conserved serine residues, with distinct functional consequences for each phosphorylation site:
Table 1: Key Phosphorylation Sites on YAP/TAZ and Their Functional Consequences
| Protein | Phosphorylation Site | Kinase | Functional Consequence |
|---|---|---|---|
| YAP | S127 | LATS1/2 | Creates 14-3-3 binding site, promotes cytoplasmic retention [38] |
| YAP | S381 | LATS1/2 | Primes for subsequent CK1δ/ε phosphorylation, leading to β-TRCP-mediated ubiquitination and degradation [38] |
| YAP | S61, S109, S164 | LATS1/2 | Additional LATS phosphorylation sites; mutation of all five sites (5SA mutant) confers oncogenic activity [38] |
| TAZ | Multiple serine residues | LATS1/2 | Similar to YAP; leads to cytoplasmic retention and degradation [37] |
The phosphorylation of YAP at Ser381 initiates a phosphodegron sequence that is subsequently phosphorylated by CK1δ/ε, creating a binding site for the SCFβ-TRCP E3 ubiquitin ligase and ultimately leading to YAP ubiquitination and proteasomal degradation [38]. This coordinated phosphorylation represents a critical mechanism for temporal regulation of YAP activity, working in concert with the spatial regulation mediated by S127 phosphorylation.
Comparative Analysis of Kinase Assay Technologies
Multiple assay formats are available for measuring kinase activity, each with distinct advantages and limitations for studying LATS1/2 and YAP/TAZ phosphorylation.
Table 2: Comparison of Major Kinase Assay Technologies
| Assay Type | Detection Principle | Pros | Cons | Best Application for Hippo Research |
|---|---|---|---|---|
| Radiometric | Measures incorporation of ³²P from [γ-³²P]ATP into substrates | High sensitivity; historically considered "gold standard" [40] | Radioactive materials; special safety and disposal requirements [40] | Initial validation studies; low-throughput mechanistic investigations |
| Antibody-Based | Phospho-specific antibodies detect phosphorylated substrates | Site-specific detection; adaptable to various formats (Western blot, ELISA) [40] | Dependent on antibody quality/availability; limited universality [40] | Specific phosphorylation site analysis (e.g., YAP pS127); validation studies |
| Luminescent (ADP-Glo) | Converts ADP to ATP, measured via luciferase luminescence | Highly sensitive; widely adopted [40] | Multi-step workflow; requires coupling enzymes; higher reagent costs [40] | Medium-throughput screening; general kinase activity assessment |
| Fluorescence-Based ADP Detection | Directly measures ADP production using fluorescence polarization (FP), TR-FRET, or intensity | Universal for any kinase; mix-and-read format; HTS compatible; supports native substrates [40] | Requires fluorescence-capable plate reader [40] | High-throughput compound screening; kinetic studies; selectivity profiling |
Assessment of Assay Technologies for MOB Complex Research
For research specifically focusing on MOB complexes in Hippo signaling, the optimal assay choice depends on the experimental goals:
Fluorescence-based ADP detection assays (e.g., Transcreener ADP²) offer significant advantages for studying MOB-regulated LATS activity due to their universality and compatibility with diverse substrate proteins [40]. This is particularly valuable when investigating how different MOB complexes (MOB1 vs. MOB4) influence LATS kinase activity toward various substrates.
Antibody-based methods remain essential for site-specific phosphorylation analysis, such as distinguishing YAP phosphorylation at S127 versus S381, which have distinct functional consequences [38]. These are invaluable for validating findings from high-throughput screens and establishing mechanistic links between MOB complex regulation and specific phosphorylation events.
Luminescent assays provide a sensitive alternative for medium-throughput applications, while radiometric assays continue to offer the highest sensitivity for specialized applications despite safety concerns [40].
Experimental Approaches for MOB Complex Studies
Direct Measurement of LATS Kinase Activity
Protocol: Fluorescence-Based LATS Kinase Assay with MOB Protein Regulation
Reagents and Materials:
- Purified LATS1 or LATS2 kinase (active form)
- MOB1 or MOB4 proteins
- ATP solution (variable concentrations)
- YAP-derived substrate peptide or full-length protein
- Fluorescence-based ADP detection kit (e.g., Transcreener ADP²)
- 384-well low-volume microplates
- Fluorescence plate reader capable of FP, TR-FRET, or FI detection
Procedure:
- Prepare kinase reaction buffer (e.g., 50 mM HEPES pH 7.5, 10 mM MgCl₂, 1 mM DTT, 0.01% Tween-20)
- Pre-incubate LATS kinase (1-10 nM) with MOB1 (activating) or MOB4 (inhibitory) proteins (10-100 nM) for 15 minutes at room temperature
- Add YAP substrate (0.1-10 μM) and initiate reaction with ATP (KM concentration ~10-50 μM)
- Incubate for 60-120 minutes at 30°C
- Stop reaction with EDTA and develop with detection reagents according to manufacturer's instructions
- Measure signal (FP, TR-FRET, or FI) and calculate ADP production as measure of kinase activity
Data Interpretation: Compare LATS kinase activity in the presence of MOB1 versus MOB4 complexes. MOB1 typically enhances LATS activity, while MOB4-containing STRIPAK complex generally suppresses it through upstream MST inhibition.
Cellular YAP/TAZ Phosphorylation Assessment
Protocol: Phospho-Specific Western Blot Analysis
Reagents and Materials:
- Cell lysates from experimental conditions
- Phospho-specific antibodies: YAP (pS127), YAP (pS381), LATS1 (pS909)
- Total YAP and LATS antibodies for normalization
- SDS-PAGE and Western blotting equipment
- ECL or fluorescence-based detection system
Procedure:
- Treat cells with experimental modulators (e.g., TRULI for LATS inhibition [39])
- Prepare cell lysates in RIPA buffer with phosphatase and protease inhibitors
- Separate proteins by SDS-PAGE (8-12% gels) and transfer to PVDF membranes
- Block membranes and incubate with primary antibodies (1:1000 dilution) overnight at 4°C
- Incubate with appropriate HRP-conjugated or fluorescent secondary antibodies
- Detect signal and quantify band intensities
- Normalize phospho-signals to total protein levels
Application for MOB Studies: This method allows researchers to assess how genetic or pharmacological manipulation of MOB complexes affects downstream YAP/TAZ phosphorylation. For example, MOB4 knockdown would be expected to increase YAP S127 phosphorylation due to enhanced MST/LATS activity, while MOB1 knockdown would decrease it.
MOB Complex-Specific Research Considerations
Distinct Regulatory Functions of MOB1 vs. MOB4
The experimental approaches for studying LATS/YAP phosphorylation must account for the fundamentally different roles of MOB complexes in Hippo signaling:
MOB1 forms an integral part of the canonical Hippo kinase cascade, directly binding to and activating LATS1/2 kinases following phosphorylation by MST1/2 [2]. Experimental strategies focusing on MOB1 should examine:
- MOB1 phosphorylation status as an indicator of upstream Hippo pathway activation
- MOB1-LATS interaction strength using co-immunoprecipitation or proximity assays
- Correlation between MOB1 expression/activation and LATS kinase activity
MOB4 functions as a core component of the STRIPAK complex, which negatively regulates Hippo signaling through phosphatase activity [9] [7]. Research approaches for MOB4 should investigate:
- STRIPAK complex assembly and its interaction with MST kinases
- MST1/2 dephosphorylation and inactivation
- Indirect effects on LATS/YAP phosphorylation through upstream regulation
Experimental Data Highlighting MOB Complex Differences
Recent studies have illuminated the distinct functions of MOB complexes:
Proteomic proximity labeling screens revealed that MOB1 and MOB4 share minimal interactors, with MOB1 predominantly associating with core Hippo components (LATS1/2, MST1/2) while MOB4 interacts primarily with STRIPAK complex members [30].
Structural studies of STRIPAK have shown that MOB4 interacts with both PP2A phosphatases and STRN proteins in a 200 × 100 × 80 Å elongated architecture, explaining its mechanism for Hippo pathway inhibition [9].
Functional experiments demonstrate that MOB4 competes with MOB1 for binding sites, providing a mechanism for fine-tuning Hippo pathway activity [7].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for Studying MOB Complex Regulation of LATS/YAP Phosphorylation
| Reagent Category | Specific Examples | Research Application | Considerations |
|---|---|---|---|
| Kinase Inhibitors | TRULI (LATS inhibitor) [39] | Chemical inhibition of LATS kinases to probe MOB complex functions | ATP-competitive; 10 μM concentration used in cellular assays |
| MST Inhibitors | XMU-MP-1 [39] | Inhibition of upstream Hippo kinases to study pathway hierarchy | May have off-target effects; use with appropriate controls |
| YAP/TEAD Inhibitors | Verteporfin [39] | Disruption of YAP-TEAD interaction to assess functional outcomes | Used at 5 μM in combination studies; validates YAP-dependent effects |
| Activity Assays | Transcreener ADP² [40] | Universal kinase activity measurement for biochemical studies | Compatible with diverse substrates; mix-and-read format |
| Phospho-Specific Antibodies | YAP (pS127), YAP (pS381), LATS1 (pS909) [38] [39] | Site-specific phosphorylation assessment in cellular contexts | Requires validation for specific applications and species |
| MOB Expression Constructs | MOB1A/B, MOB4 wild-type and mutants | Genetic manipulation of MOB complex function | Consider isoform-specific effects; MOB4 has multiple splice variants |
The choice of kinase activity assay for measuring LATS1/2 and YAP/TAZ phosphorylation downstream of MOB complexes depends heavily on the specific research question and experimental context. For high-throughput compound screening and kinetic studies, fluorescence-based ADP detection assays offer unparalleled versatility and efficiency. For validation and mechanistic studies requiring site-specific phosphorylation analysis, antibody-based methods remain essential.
Critically, researchers must account for the opposing functions of MOB1 and MOB4 complexes when designing experiments and interpreting results. MOB1 activates the Hippo kinase cascade leading to LATS-mediated YAP/TAZ phosphorylation, while MOB4-containing STRIPAK complex inhibits upstream MST kinases, thereby reducing LATS activity and YAP/TAZ phosphorylation. This fundamental distinction necessitates tailored experimental approaches for studying each MOB complex and its role in Hippo pathway regulation.
The continued development of selective small-molecule inhibitors, improved assay technologies, and more specific research tools will further enhance our understanding of how MOB complexes coordinate LATS/YAP phosphorylation in health and disease.
Visual Appendix: Signaling Pathways and Experimental Workflows
Hippo Pathway Regulation by MOB Complexes
Kinase Assay Selection Workflow
YAP Phosphorylation and Degradation Pathway
In the landscape of cellular signaling pathways, the Hippo pathway is a critical regulator of organ size, cell proliferation, and apoptosis, with its dysregulation being a hallmark of many cancers [41] [42]. Central to this pathway are the mammalian STE20-like protein kinases (MSTs) and their adaptor proteins, the Mps one binder (MOB) family. The canonical MST1–MOB1 complex acts as a tumor suppressor by activating LATS1/2 kinases, which phosphorylate and inhibit the oncogenic co-activator YAP (Yes-associated protein) [6] [41].
Emerging research has revealed a fascinating regulatory layer involving the striatin-interacting phosphatase and kinase (STRIPAK) complex. A key noncanonical complex within STRIPAK, MST4–MOB4, has been identified as a positive regulator of YAP activity, functioning in opposition to the canonical MST1–MOB1 complex [6]. In pancreatic cancer models, the MST4–MOB4 complex promotes cell growth and migration, and its expression is elevated in pancreatic cancer tissues, unlike the tumor-suppressive MST1–MOB1 [6]. This article will objectively compare the functional cellular assays used to quantify the distinct biological outcomes driven by these competing MOB complexes, providing researchers with a clear guide for experimental design.
Key Signaling Pathways: MOB1 vs. MOB4
The diagram below illustrates the core signaling dynamics between the canonical MOB1 and noncanonical MOB4 pathways, highlighting their opposing functions.
Comparative Assay Performance for Key Cellular Phenotypes
To dissect the opposing functions of MOB1 and MOB4 in the Hippo pathway, researchers employ a suite of functional cellular assays. The table below summarizes the primary assays used to quantify changes in cell proliferation, migration, and gene expression.
Table 1: Functional Assays for Quantifying MOB Complex Phenotypes
| Cellular Process | Assay Type | Key Measurable Outputs | Application in MOB1 vs. MOB4 Research |
|---|---|---|---|
| Cell Proliferation & Viability | Real-Time Cell Analysis (RTCA) [43] | Cell Index (CI); Dynamic growth curves; Half-maximal inhibitory concentration (IC50) [44] | MST4–MOB4 expression promotes CI increase; MST1–MOB1 suppresses it [6] |
| Cell Migration & Invasion | RTCA Migration Assay [43] | Migration Rate; Cell Index slope | MST4–MOB4 complex increases migration in PANC-1 cells [6] |
| Scratch Wound Healing Assay [43] | Wound Closure Rate; Semi-quantitative | Useful for initial, low-cost assessment of migratory phenotypes | |
| Gene Expression & Signaling | Single-Cell RNA Sequencing (scRNA-seq) [45] | YAP/TAZ target gene expression (e.g., CTGF, CYR61) [42] | Identifies YAP-driven transcriptomes upon MST4–MOB4 disruption |
| Immunoblot / Immunofluorescence | YAP phosphorylation status; Nuclear vs. cytoplasmic YAP localization [6] [42] | Measures Hippo pathway activity and YAP activation status |
Detailed Experimental Protocols
Real-Time Cell Analyzer (RTCA) for Proliferation and Migration
The xCELLigence RTCA system provides label-free, dynamic monitoring of cell phenotypes, overcoming the limitations of endpoint assays [43].
Proliferation/Cytotoxicity Protocol (using E-plates):
- Background Reading: Add 50 µL of culture medium to wells and perform initial measurement [43].
- Cell Seeding: Trypsinize, count cells, and seed at an optimized density (e.g., 20,000 cells/well in 200 µL medium) [43].
- Data Acquisition: Place the plate in the RTCA cradle at 37°C and 5% CO₂. Program the instrument to record impedance (displayed as Cell Index) every 5 minutes for the first 2 hours, then every 10 minutes for up to 72 hours or longer [43].
- Data Analysis: Plot Cell Index over time. A rising curve indicates proliferation. Compare the slopes and final CI values between control cells and those overexpressing MOB1 or MOB4 complexes.
Migration Protocol (using CIM-plates):
- Plate Preparation: Hydrate the upper chambers of the CIM-plate with serum-free medium in a 37°C incubator for 30 minutes.
- Cell Preparation: Serum-starve cells for 24 hours, then trypsinize and resuspend in serum-free medium.
- Loading Cells: Add 100 µL of cell suspension (e.g., 50,000 cells) to the upper chamber. Add 150 µL of medium with 10% FBS as a chemoattractant to the lower chamber.
- Data Acquisition: Monitor cell migration every 15 minutes for 24 hours. As cells migrate through the porous membrane and attach to the underside, the CI increases.
- Data Analysis: The slope of the CI curve and the final CI value are directly proportional to the rate and extent of migration, respectively.
Single-Cell RNA Sequencing to Profile YAP/TAZ Activity
scRNA-seq is powerful for uncovering the heterogeneous transcriptional programs regulated by MOB complexes [45].
Workflow for Flex Assay:
- Sample Preparation: Harvest cells (e.g., PANC-1 with modulated MST4–MOB4 or MST1–MOB1 levels). For the Flex assay, cells are fixed immediately to preserve the transcriptional state [45].
- Probe Hybridization: Incubate fixed cells with a panel of gene-specific probes targeting the whole transcriptome, including Hippo/YAP pathway targets like CTGF and CYR61 [45].
- Partitioning and Library Prep: Load the cells onto a microfluidic chip to create Gel Beads-in-Emulsions (GEMs). Within each GEM, cell barcodes are added to the transcripts. The resulting libraries are then sequenced [45].
- Data Analysis: After alignment and quality control, identify cell clusters and analyze differential gene expression. A key analysis is to compare the expression signature of YAP/TAZ target genes between cells with active MST4–MOB4 versus MST1–MOB1 signaling.
The Scientist's Toolkit: Essential Research Reagents
Successful investigation into MOB signaling requires a specific set of reagents and tools. The following table details key solutions for these functional studies.
Table 2: Key Research Reagent Solutions for MOB-Hippo Research
| Reagent / Solution | Function and Application | Example Use Case |
|---|---|---|
| Validated Antibodies | Detect protein expression, localization, and post-translational modifications. | Anti-p-YAP (Ser127) to monitor Hippo activity; Anti-MOB4 for co-immunoprecipitation [6] |
| scRNA-seq Assay Kits | Profile whole transcriptome gene expression at single-cell resolution. | 10x Genomics Flex Gene Expression Assay for fixed cells from culture or tissue [45] |
| RTCA Instrument Systems | Label-free, real-time monitoring of cell proliferation, migration, and cytotoxicity. | xCELLigence system for dynamic tracking of PANC-1 cell behavior [43] |
| MOB Expression Plasmids | Genetically manipulate the expression of MOB proteins and their mutants. | Overexpression of WT MST4 and kinase-dead (K53R) MST4 to study phosphorylation-dependent MOB4 binding [6] |
| Pathway-Specific Reporters | Visually quantify signaling pathway activity in live or fixed cells. | TEAD-luciferase reporter to measure YAP/TAZ transcriptional activity. |
Functional cellular assays are indispensable for quantifying the distinct biological outcomes of the canonical MOB1 and noncanonical STRIPAK-associated MOB4 complexes. As research reveals more complexity in Hippo pathway regulation, the precision, robustness, and dynamic data provided by assays like RTCA and scRNA-seq become ever more critical. This objective comparison of assay performance and protocols provides a framework for researchers to design rigorous experiments, ultimately accelerating our understanding of these pivotal regulatory systems in health and disease.
The Mob (Mps one binder) family of adaptor proteins represents a highly conserved group of non-catalytic proteins that serve as critical regulators of signaling pathways controlling cell proliferation, differentiation, and tissue homeostasis. Among these, MOB4 has emerged as a protein of particular interest due to its dual functionality as both a component of the STRIPAK complex (Striatin-interacting phosphatase and kinase) and a regulator of the Hippo signaling pathway [7] [8]. Unlike its canonical counterpart MOB1, which primarily functions as a core activator of the Hippo pathway, MOB4 exhibits a more complex regulatory role, often antagonizing Hippo signaling while simultaneously integrating multiple cellular inputs [7] [8]. This review provides a comprehensive comparison of two powerful in vivo model systems—Drosophila melanogaster and Danio rerio (zebrafish)—for investigating MOB4 functions in developmental processes and tissue homeostasis, with particular emphasis on their utility in STRIPAK complex research relative to canonical MOB1-Hippo signaling.
Biological Functions of MOB4 in Development and Homeostasis
MOB4 Structure and Conservation
MOB4 proteins exhibit remarkable evolutionary conservation across species, sharing approximately 80% amino acid identity and 88% similarity between Drosophila and humans [7]. Structurally, MOB4 adopts the characteristic Mob fold consisting of a four-helix bundle core with three short α-helices at the N-terminal extension, maintaining high structural homology with MOB1 despite sequence divergence in specific regions [7] [8]. This conservation underscores MOB4's fundamental biological importance and validates the use of model organisms for functional studies.
Key Functional Domains of MOB4
- STRIPAK Interaction Domain: MOB4 integrates into the STRIPAK complex through direct interactions with striatin proteins, positioning it to regulate phosphatase activity toward various kinase substrates [7] [11].
- Hippo Regulatory Region: While MOB4 does not directly activate NDR kinases like MOB1 does, it competes with MOB1 and contributes to the negative regulation of Hippo signaling through STRIPAK-mediated dephosphorylation of MST kinases [7] [8].
- Membrane Association Motifs: MOB4 participates in subcellular localization of STRIPAK complexes to various membranous structures, including the endoplasmic reticulum and nuclear envelope [46].
Table 1: Comparative Analysis of MOB4 Functions in Development and Disease
| Biological Process | MOB4 Role | Molecular Mechanism | Disease Association |
|---|---|---|---|
| Cell Proliferation | Negative regulator | STRIPAK-mediated inhibition of Hippo kinases | Various cancers |
| Neural Development | Synapse formation, neuronal migration | Cytoskeleton organization, vesicular trafficking | Neurodevelopmental disorders |
| Tissue Homeostasis | Maintenance of differentiated state | Regulation of mitochondrial function, autophagy | Age-related degeneration |
| Muscle Development | Proper muscle patterning | Regulation of myoblast fusion and differentiation | Muscular disorders |
Drosophila as a Model for MOB4 Research
Genetic Toolkit and Experimental Advantages
The fruit fly Drosophila melanogaster offers an exceptionally powerful genetic system for investigating MOB4 functions with high physiological relevance to mammalian systems. Key advantages include:
- Comprehensive Genetic Tools: The availability of GAL4/UAS system for tissue-specific manipulation, RNAi lines for knock-down studies, and CRISPR/Cas9 for precise genetic editing enables detailed functional analysis [7].
- Conserved Signaling Pathways: Core components of both STRIPAK and Hippo pathways are evolutionarily conserved in flies, with dMOB4 showing high functional similarity to human MOB4 [7] [8].
- Developmental Accessibility: External development and well-characterized embryogenesis facilitate analysis of developmental processes without invasive procedures.
- Tissue-Specific Expression Systems: Targeted genetic manipulation in specific tissues including nervous system, muscles, and epithelia allows precise dissection of MOB4 functions [47] [7].
Key Experimental Findings in Drosophila
Research in Drosophila has revealed that dMob4 is an essential gene, with null mutants failing to survive past larval stages [7]. Detailed analyses have demonstrated several critical functions:
- Neuronal Development and Function: Somatic knockdown of dMob4 results in severe neuronal defects including abnormal branching patterns, hyperbranching, incorrect synaptic connections, and disrupted microtubule organization [7]. Mutant neurons show impaired axonal transport and smaller neuromere clusters with shortened, thickened neurites.
- Synaptic Development: Drosophila Mob4 localizes to dendritic spines and regulates synapse formation through mechanisms involving endocytic membranes and vesicular trafficking [7].
- Muscle Development: While not as extensively characterized as neuronal functions, genetic interactions suggest roles in muscle patterning and function, potentially through regulation of Hippo signaling [47].
Figure 1: MOB4 Functional Interactions in Drosophila Development
Standardized Experimental Protocols for Drosophila MOB4 Research
Tissue-Specific Genetic Manipulation
- Genetic Cross Setup: Cross virgin females from tissue-specific GAL4 driver lines (e.g., elav-GAL4 for neurons, Mef2-GAL4 for muscles) with UAS-dMob4-RNAi or UAS-dMob4-overexpression males.
- Progeny Collection: Collect experimental progeny at appropriate developmental stages (embryonic, larval, or adult).
- Phenotypic Analysis: Assess developmental defects using microscopic examination, immunohistochemistry, or behavioral assays.
- Molecular Validation: Confirm manipulation efficiency via qRT-PCR, Western blotting, or RNA sequencing.
Ex Vivo Brain Culturing for Acute Interventions
The ex vivo Drosophila brain culture system enables acute pharmacological and live-imaging approaches to complement genetic studies [48]. Key steps include:
- Dissection: Isolate whole brains from third instar larvae or early pupae in sterile Schneider's insect medium.
- Culture Establishment: Transfer brains to culture chambers containing supplemented medium.
- Pharmacological Treatment: Apply specific inhibitors (e.g., kinase inhibitors, phosphatase activators) to assess acute MOB4-dependent signaling.
- Live Imaging: Monitor dynamic processes like axon guidance, synapse formation, or cytoskeletal reorganization in real-time.
This protocol is particularly valuable for studying MOB4 functions in neuronal development and allows direct manipulation of STRIPAK complex activity [48].
Zebrafish as a Model for MOB4 Research
Experimental Advantages of the Zebrafish System
Zebrafish offer complementary strengths for MOB4 research, particularly for high-resolution in vivo imaging and regenerative studies:
- Optical Clarity: Embryonic and larval transparency enables direct visualization of cellular processes in living animals [49].
- High-Fecundity: Production of hundreds of embryos weekly facilitates large-scale genetic and pharmacological screens [50] [49].
- Genetic Tractability: CRISPR/Cas9 and morpholino technologies allow efficient gene manipulation with temporal control [50].
- Regenerative Capacity: Remarkable ability to regenerate fins, heart, central nervous system, and other tissues provides unique insights into MOB4 functions in tissue homeostasis and repair [50].
Key Experimental Findings in Zebrafish
Studies in zebrafish have illuminated conserved MOB4 functions while revealing novel aspects of its regulation:
- Neurodevelopmental Roles: Mob4 knockdown in zebrafish embryos produces severe neurological defects including loss of the midbrain-hindbrain boundary, reduced eye size, and diminished head size without affecting axial trunk development [7]. These phenotypes highlight MOB4's specific requirements in brain patterning.
- Cell Proliferation Regulation: Mob4 morphants exhibit significantly reduced cell division rates in hindbrain and eye regions, indicating its critical role in controlling proliferative dynamics during development [7].
- Tissue Regeneration: While direct evidence for MOB4 in zebrafish regeneration is still emerging, its position in STRIPAK complex and connections to Hippo signaling suggest involvement in regenerative processes documented in fins, heart, and CNS [50].
Figure 2: MOB4 Functional Analysis in Zebrafish Development
Standardized Experimental Protocols for Zebrafish MOB4 Research
Morpholino-Mediated Knockdown
- Morpholino Design: Design translation-blocking or splice-blocking morpholinos targeting zfMob4 start codon or exon-intron boundaries.
- Microinjection: Inject 1-4 ng of morpholino into 1-4 cell stage zebrafish embryos using standard microinjection techniques.
- Phenotypic Scoring: Assess developmental defects at 24-48 hours post-fertilization (hpf), focusing on brain morphology, eye size, and overall growth.
- Rescue Experiments: Co-inject in vitro transcribed mob4 mRNA to confirm phenotype specificity.
In Vivo Imaging of Cellular Processes
The optical clarity of zebrafish enables unprecedented views of MOB4-dependent processes in living animals [49]:
- Transgenic Line Generation: Create mob4-promoter driven fluorescent reporter lines or fuse mob4 with fluorescent tags using CRISPR/Cas9.
- Time-Lapse Imaging: Mount live embryos in low-melting point agarose and capture high-resolution images of developing tissues over time.
- Super-Resolution Applications: Employ light-sheet or confocal microscopy to visualize subcellular localization and dynamics.
- Image Analysis: Quantify cellular behaviors including migration, division, and differentiation in mob4-manipulated embryos.
Direct Comparison: Drosophila vs. Zebrafish for MOB4 Research
Table 2: Systematic Comparison of Drosophila and Zebrafish Model Systems for MOB4 Research
| Parameter | Drosophila melanogaster | Danio rerio (Zebrafish) |
|---|---|---|
| Genetic Tractability | Extensive (GAL4/UAS, CRISPR, RNAi) | Strong (CRISPR, morpholinos, transgenesis) |
| Developmental Speed | 10 days (embryo to adult) | 3 months (embryo to sexual maturity) |
| Imaging Capabilities | Good (ex vivo and limited in vivo) | Excellent (whole organism in vivo) |
| Conservation to Human MOB4 | 80% amino acid identity | High structural and functional conservation |
| Key MOB4 Phenotypes | Larval lethality, neuronal defects, synaptic abnormalities | Brain patterning defects, reduced proliferation, small eyes |
| STRIPAK Analysis | Well-established genetic interactions | Emerging with good potential |
| Hippo Pathway Regulation | Clearly demonstrated antagonistic relationship | Likely conserved but less directly shown |
| Therapeutic Screening | Moderate-throughput possible | High-throughput chemical screening feasible |
| Regeneration Studies | Limited tissue regeneration capacity | Extensive (fin, heart, CNS, etc.) |
| Cost Considerations | Lower maintenance costs | Higher infrastructure requirements |
The STRIPAK Complex: MOB4 vs. Canonical MOB1 in Hippo Signaling
Molecular Architecture and Signaling Mechanisms
The STRIPAK complex represents a supramolecular assembly that integrates kinase and phosphatase activities to control multiple signaling pathways [51] [46] [11]. MOB4 serves as a core component of STRIPAK, while MOB1 functions as a primary activator of NDR kinases in the canonical Hippo pathway [8].
Structural organization reveals that STRIPAK complexes assemble through a "two-arm" architecture with striatin-bound PP2A as an organizing center [11]. MOB4 integrates into this complex through direct interactions, positioning it to regulate substrate accessibility and phosphorylation status of associated kinases, including Hippo pathway components.
Table 3: Functional Comparison of MOB4 vs. MOB1 in Signaling Pathways
| Characteristic | MOB4 | Canonical MOB1 |
|---|---|---|
| Primary Complex | STRIPAK | Hippo signalosome |
| NDR Kinase Binding | No stable binding | Direct activation of Warts/LATS |
| Hippo Pathway Role | Negative regulator (antagonizes) | Positive regulator (activates) |
| STRIPAK Association | Core component | Not associated |
| Conservation Level | High (80% human-fly identity) | Very high (>90% human-fly identity) |
| Developmental Functions | Neuronal development, proliferation control | Tissue growth, organ size control |
| Disease Associations | Cancer, neurological disorders | Cancer, cardiovascular diseases |
Experimental Approaches for STRIPAK-Hippo Signaling Analysis
Genetic Interaction Studies
Both Drosophila and zebrafish enable sophisticated genetic interaction analyses to delineate MOB4 functions within STRIPAK versus canonical MOB1-Hippo signaling:
- Double Mutant Analysis: Generate animals with combinations of mob4, mob1, and other STRIPAK or Hippo component mutations.
- Epistasis Tests: Determine hierarchical relationships through rescue experiments and pathway component manipulations.
- Transcriptional Readouts: Monitor Hippo pathway activity using established reporters (e.g., ex-lacZ in Drosophila, CTGF expression in zebrafish).
Biochemical Validation
Complement genetic studies with biochemical approaches to confirm molecular interactions:
- Co-Immunoprecipitation: Validate physical interactions between MOB4 and STRIPAK components versus MOB1-Hippo interactions.
- Phosphoproteomics: Identify MOB4-dependent phosphorylation changes using mass spectrometry.
- Live-Cell Imaging: Monitor complex dynamics and localization in real-time using FRET/BRET approaches.
Figure 3: MOB4-STRIPAK vs. MOB1-Hippo Signaling Relationships
Research Reagent Solutions for MOB4 Studies
Table 4: Essential Research Reagents for MOB4 Investigation in Model Systems
| Reagent Category | Specific Examples | Research Applications | Availability |
|---|---|---|---|
| Genetic Tools | UAS-dMob4-RNAi (Drosophila), mob4 morpholinos (Zebrafish) | Loss-of-function studies | Public stock centers, commercial suppliers |
| Expression Constructs | UAS-dMob4, CMV-zfMob4-GFP | Overexpression, localization | cDNA libraries, gene synthesis |
| Antibodies | Anti-MOB4, Anti-STRIPAK components | Protein detection, localization | Commercial, custom generation |
| Reporter Lines | Hippo pathway reporters (ex-lacZ, CTGF-GFP) | Pathway activity monitoring | Published designs, custom generation |
| Pharmacological Agents | Kinase inhibitors, phosphatase activators | Acute pathway manipulation | Commercial suppliers |
| Imaging Tools | mob4-promoter:GFP, tagged MOB4 lines | Live imaging, dynamics | CRISPR/Cas9 generation |
Drosophila and zebrafish provide complementary model systems for elucidating MOB4 functions in development and tissue homeostasis, each offering unique experimental advantages. Drosophila excels in genetic dissection of MOB4 roles in neuronal development and complex tissue interactions, while zebrafish offers unparalleled capabilities for in vivo visualization and regeneration studies. Both systems consistently demonstrate MOB4's essential functions in regulation of cell proliferation, neural development, and tissue patterning, primarily through its integration into the STRIPAK complex and modulation of Hippo signaling.
Future research directions should leverage the strengths of both systems to address outstanding questions, including tissue-specific requirements for MOB4, its roles in adult homeostasis and aging, and potential contributions to disease processes. The continued development of more sophisticated genetic tools, imaging technologies, and biochemical approaches in both Drosophila and zebrafish will undoubtedly accelerate our understanding of this critical regulatory protein and its connections to human development and disease.
Resolving Experimental Challenges in Differentiating MOB1 and MOB4 Signaling
The Hippo signaling pathway is a central regulator of tissue homeostasis, cell proliferation, and apoptosis, with its dysregulation implicated in various cancers and other diseases. At the core of this pathway are the MST kinases and MOB adapter proteins, which form critical complexes that regulate downstream effectors. However, the existence of multiple MST and MOB family members presents a significant challenge for genetic perturbation studies, as functional redundancy and compensatory mechanisms can obscure phenotypic effects and lead to misinterpretation of experimental results. This guide systematically compares the distinct and overlapping functions of key MST and MOB family members, with particular focus on the STRIPAK complex MOB4 versus canonical MOB1 Hippo signaling, providing strategies to address redundancy in experimental designs.
Table 1: Core Components of MST Kinase and MOB Protein Families
| Protein Family | Key Members | Primary Binding Partners | Cellular Function | Pathway Association |
|---|---|---|---|---|
| MST Kinases | MST1, MST2 | MOB1 | Pro-apoptotic, tumor suppressor | Canonical Hippo |
| MST4 | MOB4 | Promotes cell migration/growth | STRIPAK Complex | |
| MOB Proteins | MOB1A, MOB1B | MST1/2, LATS1/2 | Tumor suppressor, NDR kinase activator | Canonical Hippo |
| MOB2 | NDR1/2 | Inhibits cell migration | Hippo-like | |
| MOB4 (Phocein) | MST4, STRNs | Regulates cell polarity, antagonizes Hippo | STRIPAK Complex |
Comparative Biology of MOB1 and MOB4 Complexes
Canonical MOB1 Signaling
MOB1 proteins (MOB1A and MOB1B) serve as essential adapters in the canonical Hippo pathway, functioning as tumor suppressors that regulate tissue homeostasis. When activated, MST1/2 kinases phosphorylate and activate MOB1, which subsequently interacts with and fully activates LATS1/2 kinases. This activation cascade leads to the phosphorylation and cytoplasmic retention of the transcriptional coactivators YAP/TAZ, thereby inhibiting their pro-growth transcriptional programs [41] [8]. The MST1-MOB1 complex represents a fundamental growth-restraining module that is frequently dysregulated in human cancers. MOB1's role as a tumor suppressor is evolutionarily conserved, with studies in Drosophila demonstrating that Mob1 (Mats) controls cell proliferation and apoptosis through interaction with LATS (Warts) [52].
Non-Canonical MOB4 Signaling
In contrast to MOB1, MOB4 (also known as Phocein) forms a complex with MST4 kinase and functions as a component of the STRIPAK complex (Striatin-Interacting Phosphatase and Kinase). Surprisingly, despite structural similarities to the MST1-MOB1 complex, the MST4-MOB4 complex exhibits opposite biological functions. Rather than suppressing growth, MST4-MOB4 promotes cell proliferation and migration in cancers such as pancreatic cancer [6]. The MST4-MOB4 complex disrupts the tumor-suppressive MST1-MOB1 complex through alternative pairing, thereby increasing YAP activity and driving oncogenic progression [6]. This functional antagonism represents a critical node of regulation within the broader Hippo signaling network.
Structural Basis for Functional Diversification
Although the overall structure of the MST4-MOB4 complex resembles that of MST1-MOB1, evolutionary divergence of key interface residues enables distinct functional properties. MST4 interacts with MOB4 in a phosphorylation-dependent manner that requires autophosphorylation at Thr-327/328 within its linker region [6]. This specific interaction mode allows MST4-MOB4 to compete with and disrupt MST1-MOB1 complex formation, creating a regulatory switch that controls the output of Hippo signaling. The structural conservation yet functional opposition between these complexes highlights the importance of comprehensive approaches when targeting these pathways experimentally.
Experimental Approaches and Knockdown Strategies
Addressing Compensation in Genetic Perturbations
The high degree of homology between MST and MOB family members necessitates strategic approaches to genetic perturbation to overcome compensatory mechanisms:
Combined Knockdown Strategies: Simultaneous targeting of multiple family members is often required to observe clear phenotypes. For example, studies of MOB proteins have revealed that complete functional ablation may require targeting multiple MOB genes concurrently due to overlapping functions [8] [52].
Isoform-Specific Reagents: When targeting specific MOB family members, ensure reagents are designed to avoid cross-reactivity. The four MOB classes (MOB1, MOB2, MOB3, and MOB4) share conserved domains but have distinct interaction profiles [8].
Validation of Knockdown Efficiency: Use multiple antibodies to confirm successful protein depletion, as commercial antibodies may vary in specificity for different MOB family members.
MST1 Targeting Methodologies
Successful MST1 knockdown has been achieved using both siRNA and shRNA approaches:
In Vitro siRNA Protocol: Transfert chondrocytes with 50 nM of MST1-targeting siRNA duplexes using Lipofectamine 3000 when cells reach 60-70% confluence. Incubate for 3 hours before replacing with fresh medium and conducting subsequent analyses after 48-72 hours [53].
In Vivo shRNA Approach: Utilize lentiviral vectors encoding MST1-specific shRNA for stable knockdown in animal models. For osteoarthritis studies, this approach successfully delayed disease progression by reducing inflammation, ECM degradation, and apoptosis while enhancing mitophagy [53].
CRISPR/Cas9 Knockout: For complete MST1 ablation, CRISPR/Cas9 systems can be employed. Mst1 knockout mice have been used to study diabetic cardiomyopathy, revealing MST1's role in regulating mitochondrial fission through Drp1 activation [54].
MOB Protein Targeting Approaches
Distinct strategies are required for different MOB family members due to their diverse functions:
MOB1 Knockdown: Target both MOB1A and MOB1B isoforms simultaneously to completely disrupt canonical Hippo signaling. MOB1 knockdown leads to YAP/TAZ activation and promotes cell proliferation.
MOB2 Targeting: Employ CRISPR/Cas9 with sgRNA (5′-AGAAGCCCGCTGCGGAGGAG-3′) for MOB2 knockout. MOB2 knockout promotes migration and invasion in hepatocellular carcinoma cells by enhancing YAP activity [55].
MOB4 Perturbation: Use specific shRNAs to target MOB4 without affecting other MOB family members. MOB4 knockdown impairs cancer cell growth and migration by disrupting the MST4-MOB4 complex [6].
Table 2: Knockdown/Knockout Outcomes for MST and MOB Proteins
| Target Protein | Experimental Model | Primary Phenotypic Outcomes | Compensatory Mechanisms Observed |
|---|---|---|---|
| MST1 | Human chondrocytes (siRNA) | Reduced inflammation, ECM degradation, apoptosis; enhanced mitophagy | Potential compensation by MST2 requires verification |
| MST1 | Mouse diabetic cardiomyopathy model (KO) | Improved cardiac function, reduced mitochondrial fission | Comprehensive cardiac evaluation needed |
| MOB2 | Human HCC cells (CRISPR/Cas9) | Enhanced cell migration and invasion; increased YAP activity | Assessment of other MOB proteins recommended |
| MOB4 | Pancreatic cancer cells | Inhibited cell growth and migration; reduced YAP activity | MST1-MOB1 complex activity should be monitored |
| Combined MOB1/4 | Theoretical approach | Complete disruption of both canonical and STRIPAK-Hippo signaling | Requires careful titration to avoid toxicity |
Research Reagent Solutions
Essential reagents for studying MST kinases and MOB proteins include:
MST1 Antibodies: For Western blot (1:1,000 dilution) and immunofluorescence to validate knockdown efficiency and assess subcellular localization [53] [54].
MOB Class-Specific Antibodies: Crucial for distinguishing between MOB1, MOB2, and MOB4 proteins, as they may have overlapping migration patterns [8] [52].
Lentiviral Constructs: For stable shRNA delivery (e.g., lentiCRISPRv2 for CRISPR/Cas9); use puromycin selection (1.0 μg/mL) for 2 weeks to establish stable cell lines [55] [6].
Pathway Reporters: YAP/TAZ localization assays and TEAD-luciferase reporters to monitor functional outcomes of perturbations.
Mitochondrial Isolation Kits: For assessing mitochondrial function and Drp1 translocation following MST1 perturbation [54].
Signaling Pathway Diagrams
Diagram 1: STRIPAK-MOB4 Antagonizes Canonical MOB1 Hippo Signaling. The MST4-MOB4 complex disrupts tumor-suppressive MST1-MOB1 signaling, leading to YAP/TAZ activation.
Discussion and Future Perspectives
The complex interplay between different MST kinases and MOB proteins necessitates carefully designed experimental approaches to overcome redundancy and compensation. The opposing functions of canonical MOB1 and STRIPAK-associated MOB4 highlight the sophisticated regulatory balance controlling Hippo pathway output. Successful targeting of these complexes requires:
- Combinatorial approaches that simultaneously address multiple family members
- Validation strategies that confirm specific protein depletion without compensatory upregulation of paralogs
- Functional assays that measure pathway activity beyond simple protein expression
Future research should focus on developing more specific inhibitors and activators of these complexes, as well as sophisticated genetic models that enable spatiotemporal control of MST and MOB protein expression. The emerging understanding of STRIPAK complex regulation of Hippo signaling opens new therapeutic opportunities for cancer and other diseases characterized by dysregulated tissue growth.
Optimizing Conditions for Phosphorylation-Dependent Interaction Studies
The Mps One Binder (MOB) proteins are evolutionarily conserved eukaryotic regulators of kinase signaling, with pivotal roles in cell proliferation, morphogenesis, and exit from the cell cycle [32]. Within the human MOB family, canonical MOB1 is a well-characterized integrator of the Hippo tumor suppressor pathway, while MOB4 represents a less understood non-canonical member whose functions appear distinct from Hippo signaling [32]. A precise comparison of their phosphorylation-dependent interactions is critical for advancing fundamental research and drug discovery, as the Hippo pathway is a major regulator of organ size and its dysregulation is a hallmark of many cancers [41] [56].
This guide provides a structured experimental framework for comparing MOB1 and MOB4, focusing on the optimization of conditions for studying their phosphorylation-dependent interactomes. We objectively compare known interaction partners, structural features, and functional outputs by synthesizing data from recent interaction proteomics, biochemical, and biophysical studies. The protocols and data presented are designed to equip researchers with the methodologies needed to delineate the unique signaling landscapes governed by these two regulatory proteins.
MOB Protein Family: Canonical MOB1 vs. Non-Canonical MOB4
The mammalian MOB family comprises seven members, grouped into four subfamilies, each with orthology to distinct Drosophila MOB proteins [32]. MOB1A and MOB1B (here collectively MOB1) are the canonical members, best known for their role as core components and activators of the Hippo pathway kinases LATS1/2 and NDR1/2 [32] [41]. In contrast, MOB3A, MOB3B, MOB3C, and MOB4 form a separate group that does not associate with LATS or NDR kinases, indicating a functional divergence [32].
Table 1: Core Characteristics of MOB1 and MOB4
| Feature | MOB1 (Canonical) | MOB4 (Non-Canonical) |
|---|---|---|
| Key Signaling Pathway | Hippo, NDR, Mitotic Exit Network [32] | Not associated with core Hippo signaling; specific pathways are an active area of research [32] |
| Established Kinase Partners | MST1/2 (upstream), LATS1/2, NDR1/2 (downstream) [32] | Does not appear to associate with LATS or NDR kinases [32] |
| Phospho-Regulation | Phosphorylated by MST1/2 at T12 and T35; autoregulates binding to upstream and downstream partners [32] | Largely unknown; presumed to be regulated by distinct mechanisms |
| Cellular Functions | Cell proliferation, apoptosis, contact inhibition, organ size control [41] [56] | Less defined; proposed roles in neuronal function and other specialized contexts |
Experimental Comparison of MOB1 and MOB4 Interactions
A side-by-side comparison of interaction profiles and biochemical properties is essential for differentiating MOB1 and MOB4 functions. The following data, derived from interaction proteomics and quantitative biochemical studies, provides a foundation for this comparison.
Interaction Partner Profiles
MOB1 engages with multiple protein complexes in a phosphorylation-dependent manner. Beyond its well-known role in bridging MST1/2 to LATS1/2, interaction proteomics has revealed that MOB1 also associates with the PP6 phosphatase complex and a module containing the Rho guanine exchange factors DOCK6-8 [32]. The binding to MST1/2 is direct and utilizes MOB1's phosphopeptide-binding domain, whereas the recruitment mechanisms for PP6 and DOCK complexes show key differences, suggesting alternative binding modes [32]. In contrast, the interaction network for MOB4 remains largely uncharacterized, representing a significant knowledge gap.
Table 2: Experimentally Determined Interaction Profiles
| Parameter | MOB1 | MOB4 |
|---|---|---|
| Upstream Kinase Partners | MST1, MST2 (confirmed) [32] | Limited data |
| Downstream Kinase Partners | LATS1, LATS2, NDR1, NDR2 (confirmed) [32] | No association with LATS/NDR kinases [32] |
| Other Signaling Complexes | PP6 phosphatase complex, DOCK6-8 GEF complex [32] | Limited data |
| Structural Basis of Binding | Characterized: N-term phosphosites regulate LATS/NDR binding; central domain binds MST phosphopeptides [32] | Limited data |
Quantitative Binding Affinities and Phospho-Regulation
The affinity of MOB1 for its partners is tightly regulated by phosphorylation. Quantitative biochemical studies show that phosphorylation of MOB1 at N-terminal sites (e.g., T12, T35) differentially regulates its interaction with upstream (MST) versus downstream (LATS) kinases [32]. For instance, phospho-mimetic mutations can enhance binding to downstream effectors while potentially weakening interactions with upstream regulators. The binding affinity of MOB1 for a phosphorylated MST2 peptide (pT378) has been structurally validated [32]. Similar quantitative data for MOB4 is not currently available in the scientific literature.
Table 3: Quantitative Biochemical Data for MOB1
| Interaction | Affinity (KD)/Notes | Phospho-Regulation |
|---|---|---|
| MOB1 : MST2 (pT378) | Structurally characterized phosphopeptide-binding interaction [32] | Phospho-dependent (MST2 phosphorylation required) |
| MOB1 : LATS1 | Enhanced by MOB1 N-terminal phosphorylation (e.g., T12, T35) [32] | Phospho-dependent (MOB1 phosphorylation promotes) |
| MOB1 : PP6 Complex | Recruited via mechanism distinct from MST1/2 binding [32] | Phospho-dependent (specific mechanism under investigation) |
Essential Protocols for Phosphorylation-Dependent Interaction Studies
Protocol 1: Interaction Proteomics for MOB Complexes
This protocol is adapted from studies identifying MOB1-associated complexes and can be applied to profile MOB4 interactions [32].
- Plasmid Construction: Generate plasmids for expressing tagged MOB proteins (e.g., MOB1A, MOB4). Use a modified pETM-30 vector for E. coli expression to produce N-terminal dual 6xhistidine (HIS) and glutathione S-transferase (GST) tagged proteins with a cleavable Tobacco Etch Virus (TEV) protease site [32].
- Recombinant Protein Expression and Purification:
- Express proteins in E. coli BL21 (DE3) CodonPlus RIL cells.
- Purify proteins in batch using glutathione-Sepharose resin.
- Elute by cleaving the affinity tags with HIS-tagged TEV protease.
- Remove TEV protease via subtractive immobilized-metal affinity chromatography.
- Perform final purification and buffer exchange using size exclusion chromatography (SEC) with a Superdex 75 column [32].
- Cell Culture and Transfection: Culture relevant mammalian cell lines (e.g., HEK293T). Transfect with plasmids encoding tagged MOB constructs (wild-type, phospho-mimetic, phospho-deficient).
- Affinity Purification and Mass Spectrometry:
- Lyse cells in a suitable buffer (e.g., 50 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.5% NP-40, protease and phosphatase inhibitors).
- Incubate lysates with glutathione-Sepharose beads to capture GST-tagged MOB complexes.
- Wash beads extensively, then elute complexes.
- Digest eluted proteins with trypsin and analyze peptides by liquid chromatography-tandem mass spectrometry (LC-MS/MS) [32].
- Data Analysis: Identify MOB-specific interactors by comparing against control purifications. Use quantitative proteomics (e.g., iTRAQ, TMT) to assess phosphorylation-dependent changes in the interactome [57].
Protocol 2: Assessing STRIPAK-Dependent Phosphorylation
The STRIPAK complex is a key regulator of phosphorylation in multiple pathways. This protocol uses global phosphoproteomics to identify its targets, a method applicable for studying MOB regulation [57].
- Strain Generation: Use wild-type and STRIPAK mutant strains (e.g., Δpro11, Δpro11Δpro22, Δpp2Ac1Δpro22) from a model system like the filamentous fungus Sordaria macrospora [57] [58].
- Protein Extraction and Digestion: Grow strains for 3 days in liquid culture. Isolate protein extracts and digest with trypsin.
- Phosphopeptide Enrichment: Enrich phosphopeptides from the digested samples using TiO2 enrichment followed by Hydrophilic Interaction Liquid Chromatography (HILIC) [57].
- LC-MS/MS and Data Analysis:
- Analyze enriched phosphopeptides by nano-HPLC and MS/MS.
- Identify and quantify phosphorylation sites across wild-type and mutant strains.
- Proteins with significantly altered phosphorylation in STRIPAK mutants are considered putative direct or indirect targets. In a recent study, this method identified 129 such proteins, including the RNA-binding protein GUL1 [57].
The Scientist's Toolkit: Key Research Reagents
Table 4: Essential Reagents for MOB Protein Research
| Reagent / Resource | Function / Application | Example / Source |
|---|---|---|
| MOB Expression Plasmids | Expression of wild-type, phospho-mimetic (e.g., Asp, Glu), and phospho-deficient (e.g., Ala) mutants for functional studies. | pETM-30 vector system [32] |
| Specific Antibodies | Detection of proteins and post-translational modifications via immunoblotting, immunoprecipitation. | Anti-phospho-MOB1 (T12, T35) [32] |
| Kinase Assay Kits | In vitro measurement of kinase activity for partners like LATS1/2 and MST1/2. | Commercial kits from vendors like Cell Signaling Technology |
| Hippo Pathway Inhibitors/Activators | Pharmacological modulation of pathway activity to study functional outcomes. | VT107 (Hippo inhibitor) [59]; Verteporfin (YAP-TEAD inhibitor) [41] |
| Proteomics & Phosphoproteomics Platforms | Large-scale, unbiased identification of interaction partners and phosphorylation sites. | iTRAQ-based quantification; TiO2 phosphopeptide enrichment [57] |
Signaling Pathway Diagrams
Diagram Title: MOB Protein Signaling Networks
This diagram illustrates the established canonical MOB1 pathway integrated with the Hippo signaling cascade and its regulation by the STRIPAK complex, contrasted with the less-defined signaling context of non-canonical MOB4.
Diagram Title: Workflow for MOB Interaction Studies
This workflow outlines the key stages for a comprehensive study of phosphorylation-dependent MOB interactions, from molecular cloning and proteomics to functional validation.
A rigorous, multi-faceted approach is essential for optimizing the study of MOB1 and MOB4 interactions. As detailed in this guide, leveraging interaction proteomics and global phosphoproteomics provides an unbiased map of binding partners and their regulation. Following this with quantitative biochemical and biophysical assays is crucial to measure the strength and dynamics of these interactions. Finally, the use of phospho-specific mutants in cellular models allows researchers to define the functional consequences of phosphorylation in a physiological context.
While the role of MOB1 in the Hippo pathway is increasingly well-understood, the functional landscape of MOB4 remains a fertile ground for discovery. The experimental frameworks and comparisons provided here are designed to accelerate research into both proteins, ultimately contributing to a more complete understanding of their roles in health and disease, particularly in cancer biology where Hippo signaling is frequently dysregulated.
Disentangling STRIPAK-Specific Functions from Alternative MOB4 Complexes
The monopolar spindle-one-binder (MOB) family of adaptor proteins represents crucial regulatory hubs in cellular signaling, with MOB4 emerging as a particularly compelling subject of study due to its dual affiliation with both canonical and non-canonical protein complexes. As a highly conserved non-catalytic scaffold, MOB4 participates in fundamental processes including cell proliferation, tissue development, and tumorigenesis, yet its distinct functions within different macromolecular assemblies remain incompletely characterized [7]. This analysis systematically disentangles MOB4-specific functions by comparing its role within the Striatin-Interacting Phosphatase and Kinase (STRIPAK) complex against its functions in alternative complexes, particularly with Mammalian STE20-like kinase 4 (MST4).
The STRIPAK complex is a massive supramolecular assembly that integrates both phosphatase (PP2A) and kinase activities, serving as an evolutionary conserved signaling hub across eukaryotic organisms [60] [11]. Within this complex, MOB4 functions as a core structural component, with recent cryo-EM studies revealing its precise positioning within the complex architecture [9]. Simultaneously, MOB4 forms a distinct complex with MST4 that operates outside the STRIPAK assembly, engaging in direct regulatory crosstalk with the Hippo signaling pathway [6]. This comparison guide provides an objective analysis of these MOB4 complexes, presenting structural, functional, and experimental data to equip researchers with methodologies for distinguishing STRIPAK-specific MOB4 functions from its alternative partnerships in both physiological and pathological contexts.
Structural Organization: Distinct Architectures of MOB4 Complexes
STRIPAK Complex Architecture
The STRIPAK complex represents a non-canonical protein phosphatase 2A (PP2A) holoenzyme characterized by an elaborate architecture that deviates fundamentally from traditional PP2A complexes. Recent cryo-EM structural analysis of the human STRIPAK core complex at 3.2 Å resolution has illuminated its unique composition and organization [9]. Unlike trimeric PP2A holoenzymes, STRIPAK incorporates four copies of the regulatory subunit STRN3 (striatin-3), which form an elongated homotetrameric scaffold through their coiled-coil domains. This STRN3 tetramer serves as a central platform that organizes one copy each of the PP2A catalytic core (PP2AA-PP2AC heterodimer), STRIP1, and MOB4 into an asymmetric assembly measuring approximately 200 × 100 × 80 Å [9].
Within this architecture, MOB4 occupies a critical positioning at one end of the complex, where it interfaces with both STRIP1 and the WD40 domain of STRN3, creating a connection point that contributes to overall complex stability. The structural analysis further revealed an inositol hexakisphosphate (IP6) molecule acting as a structural cofactor for STRIP1, highlighting the sophisticated organization of this massive phosphatase complex [9]. This elaborate architecture enables STRIPAK to function as an integrated signaling node that coordinates phosphatase and kinase activities within the Hippo pathway and beyond.
MST4-MOB4 Complex Architecture
In contrast to the massive STRIPAK assembly, the alternative MST4-MOB4 complex presents a considerably simpler binary structure. Biochemical and structural analyses demonstrate that MST4 directly interacts with MOB4 in a phosphorylation-dependent manner to form a stable heterodimeric complex [6]. Despite its simpler composition, the MST4-MOB4 complex exhibits remarkable structural resemblance to the canonical MST1-MOB1 complex of the Hippo pathway, suggesting evolutionary conservation of this kinase-adaptor pairing mechanism [6].
The interaction interface between MST4 and MOB4 requires autophosphorylation of MST4 at Thr-327/328 residues within its linker region, with the kinase-inactive MST4 (MST4-K53R) failing to bind MOB4 [6]. Pulldown assays using truncated MST4 variants identified amino acids 316-335 as the minimal region essential for MOB4 binding, highlighting the critical importance of this phosphorylated segment for complex formation [6]. The MST4-MOB4 complex exhibits a dissociation constant (Kd) of 1.67 μM as measured by bio-layer interferometry, indicating a specific and moderately strong interaction [6].
Table 1: Structural Comparison of MOB4-Containing Complexes
| Structural Feature | STRIPAK Complex | MST4-MOB4 Complex |
|---|---|---|
| Overall Architecture | Multimeric (≥8 subunits) | Binary heterodimer |
| Core Components | PP2AA, PP2AC, STRN3 (×4), STRIP1, MOB4 | MST4, MOB4 |
| MOB4 Binding Partners | STRIP1, STRN3 WD40 domain | MST4 kinase domain |
| Structural Basis | Cryo-EM structure resolved at 3.2 Å | Resembles MST1-MOB1 complex structure |
| Stoichiometry | Asymmetric (1 MOB4: 4 STRN3) | 1:1 MST4:MOB4 ratio |
| Key Structural Elements | STRN3 coiled-coil tetramer, IP6 cofactor | MST4 linker region (phospho-Thr-327/328) |
Functional Consequences: Signaling Outputs and Biological Effects
STRIPAK-MOB4 Functions in Hippo Signaling and Neurodevelopment
The STRIPAK complex functions as a master regulator of Hippo signaling through its negative regulation of MST1/2 kinases, the core kinases in the Hippo pathway cascade. STRIPAK-mediated dephosphorylation of MST1/2 at their activation loops (T180 for MST2) maintains these kinases in an inhibited state, thereby preventing phosphorylation of downstream effectors LATS1/2 and ultimately leading to YAP/TAZ activation [11] [9]. Within this regulatory framework, MOB4 serves as an integral structural component that contributes to complex assembly and function rather than directly engaging catalytic activities.
Beyond Hippo regulation, STRIPAK-MOB4 plays essential roles in neuronal development and function. MOB4 demonstrates predominant expression in the central nervous system, particularly localizing to dendritic spines and exhibiting strict somato-dendritic distribution without axonal presence [7]. Genetic studies in Drosophila reveal that MOB4 deficiency causes severe neurodevelopmental defects, including abnormal branching patterns, hyperbranching, disrupted microtubule organization, and impaired synaptic development [7]. Similarly, zebrafish morphants with MOB4 knockdown exhibit diminished midbrain-hindbrain boundaries, reduced eye size, and impaired cell division in neural tissues [7]. These neurological functions align with the original identification of MOB4 (initially termed phocein) as a interactor of striatins in neural tissues [7].
Recent research has further elucidated STRIPAK-MOB4 functions in peripheral nervous system development, particularly in Schwann cell-mediated radial sorting of axons. The complex interacts with active RAC1-GTP to regulate cytoskeletal reorganization and lamellipodia formation, while simultaneously controlling YAP/TAZ activity and laminin receptor expression essential for proper myelination [12].
MST4-MOB4 Oncogenic Functions in Hippo Pathway Regulation
The MST4-MOB4 complex operates as a positive regulator of YAP activity through a distinct mechanism that directly counteracts the canonical Hippo pathway. Unlike the tumor-suppressive MST1-MOB1 complex, MST4-MOB4 exhibits potent oncogenic properties, particularly in pancreatic cancer models [6]. This complex promotes cell growth and migration in PANC-1 pancreatic cancer cells, with elevated expression levels of both MST4 and MOB4 correlating positively in pancreatic cancer tissues while MST1 expression is concurrently down-regulated [6].
The mechanistic basis for this functional antagonism lies in the competitive binding behavior of MOB4. Due to divergent evolution of key interface residues, MOB4 preferentially binds MST4 over MST1, thereby disrupting formation of the tumor-suppressive MST1-MOB1 complex through alternative pairing [6]. This molecular competition effectively increases YAP nuclear localization and transcriptional activity by reducing LATS1/2 kinase activation, establishing the MST4-MOB4 complex as a noncanonical regulator of Hippo-YAP signaling with oncogenic potential [6].
Table 2: Functional Comparison of MOB4 Complexes in Hippo Signaling
| Functional Aspect | STRIPAK-MOB4 Complex | MST4-MOB4 Complex |
|---|---|---|
| Hippo Pathway Regulation | Negative regulator of MST1/2 | Positive regulator of YAP |
| Effect on YAP/TAZ | Indirect activation via MST1/2 inhibition | Direct activation via MST1-MOB1 disruption |
| Biological Role in Cancer | Context-dependent; can be tumor-promoting | Oncogenic (e.g., pancreatic cancer) |
| Cellular Phenotypes | Regulates cell polarity, migration | Promotes proliferation, migration |
| Neuronal Functions | Essential for neurodevelopment, synapse formation | Less characterized in neuronal contexts |
| Developmental Roles | Peripheral nerve development, radial sorting | Not well established |
Experimental Approaches: Methodologies for Distinguishing MOB4 Complexes
Structural and Biochemical Characterization Techniques
Co-immunoprecipitation (Co-IP) and Pulldown Assays: These fundamental techniques allow researchers to distinguish MOB4 complex constituents under different physiological conditions. For STRIPAK identification, immunoprecipitation of core components like STRN3 or STRIP1 should co-precipitate MOB4 along with PP2A subunits [11] [9]. Conversely, MST4 immunoprecipitation specifically captures MOB4 without STRIPAK components in the alternative complex [6]. Critical methodological considerations include:
- Using phosphorylation-preserving conditions (phosphatase inhibitors) for MST4-MOB4 interactions
- Employing crosslinking approaches to stabilize transient interactions
- Combining with phosphorylation status analysis of MST kinases
In Vitro Reconstitution and Binding Affinity Measurements: Direct protein-protein interactions between MOB4 and its partners can be quantified using purified recombinant proteins. MST4-MOB4 binding exhibits a Kd of 1.67 μM as measured by bio-layer interferometry [6]. For STRIPAK, structural studies have utilized the biGBac expression system to reconstitute the entire core complex (PP2AA/C, STRN3, STRIP1, MOB4) for biochemical and structural analyses [9]. Key parameters include:
- Testing phosphorylation-dependence through λ-phosphatase treatment
- Using kinase-inactive mutants (MST4-K53R) as negative controls
- Employing size exclusion chromatography to monitor complex formation
Advanced Structural Biology Approaches: Cryo-electron microscopy has proven indispensable for determining the architecture of massive complexes like STRIPAK. The 3.2 Å structure of the human STRIPAK core revealed the unexpected STRN3 tetramer and MOB4 positioning [9]. For the smaller MST4-MOB4 complex, crystallography provides atomic-level details of the interface, revealing structural conservation with MST1-MOB1 despite functional divergence [6].
Functional and Cellular Assays
Hippo Signaling Pathway Reporter Assays: YAP/TAZ transcriptional activity serves as a key functional readout for differentiating MOB4 complex functions. STRIPAK-MOB4 typically increases YAP activity through MST1/2 inhibition, while MST4-MOB4 enhances YAP activity through competitive disruption of MST1-MOB1 [6] [11]. Standardized methodologies include:
- TEAD-luciferase reporter constructs for YAP/TAZ activity
- Monitoring YAP/TAZ localization via immunofluorescence
- Quantitative RT-PCR for YAP target genes (CTGF, CYR61)
Cell Phenotyping in Loss-of-Function and Gain-of-Function Models: Genetic manipulation approaches effectively distinguish complex-specific functions. STRIPAK component depletion (SIKE1, SLMAP, STRN3, STRIP1) increases MST2 activation and YAP phosphorylation, whereas MST4-MOB4 overexpression promotes proliferation and migration in pancreatic cancer cells [6] [11]. Recommended approaches include:
- CRISPR/Cas9 knockout of specific components
- siRNA-mediated targeted protein depletion
- Inducible expression systems for gain-of-function studies
- Migration assays (wound healing, Transwell) for functional characterization
Proximity-Dependent Biotin Identification (BioID): This powerful proximity-labeling technique enables unbiased mapping of MOB4 protein interactomes under near-physiological conditions. BioID has identified over 200 potential MOB protein interactions, with MOB4 specifically associating with STRIPAK components [30]. Key methodological elements include:
- N-terminal BirA*-tagging of MOB4 to avoid functional disruption
- Tetracycline-inducible expression systems for controlled biotinylation
- Quantitative mass spectrometry for interaction quantification
- Comparative analysis across multiple MOB family members
Visualization of Signaling Pathways and Experimental Workflows
Diagram 1: MOB4 Complex Regulation of Hippo Signaling. The STRIPAK-MOB4 complex inhibits MST1-MOB1, while the MST4-MOB4 complex directly disrupts MST1-MOB1 formation. Both mechanisms lead to LATS1/2 inhibition and YAP/TAZ activation, promoting cell growth and migration.
Diagram 2: Experimental Workflow for Distinguishing MOB4 Complexes. Integrated approach combining biochemical, proteomic, structural, and functional methods to differentiate between STRIPAK-MOB4 and MST4-MOB4 complexes.
Research Reagent Solutions: Essential Tools for MOB4 Complex Studies
Table 3: Key Research Reagents for MOB4 Complex Investigation
| Reagent Category | Specific Examples | Research Application | Considerations |
|---|---|---|---|
| Expression Constructs | Full-length MOB4 isoforms, MST4 (WT/K53R), STRN3 fragments, STRIP1 | Complex reconstitution, interaction mapping | Include phosphorylation-competent and deficient mutants |
| Cell Lines | HEK293T, HeLa Flp-In T-REx, PANC-1 pancreatic cancer cells | Interaction studies, pathway analysis, functional assays | Select based on endogenous MOB4 expression and context |
| Antibodies | Anti-MOB4, anti-MST4, anti-STRN3, anti-phospho-MST1/2 (T183/T180) | Immunoprecipitation, Western blot, localization | Validate specificity for intended applications |
| Proteomic Tools | BirA*-MOB4 fusion constructs, streptavidin beads, quantitative MS | Proximity labeling, interactome mapping | Use inducible systems to control biotinylation timing |
| Structural Biology Reagents | biGBac system for complex expression, cryo-EM grids | Structural determination of complexes | Optimize purification protocols for complex stability |
| Functional Assay Tools | TEAD-luciferase reporters, Matrigel for migration assays | YAP/TAZ activity, cell phenotyping | Include appropriate controls for pathway specificity |
This comparative analysis elucidates the dual existence of MOB4 within two structurally and functionally distinct complexes that converge on Hippo pathway regulation while mediating diverse biological outputs. The STRIPAK-MOB4 complex represents an integrated phosphatase-kinase signaling hub that negatively regulates MST1/2 through its multimeric architecture, playing essential roles in neurodevelopment and cellular homeostasis. In contrast, the MST4-MOB4 binary complex functions as an oncogenic regulator that promotes YAP activation through competitive disruption of the tumor-suppressive MST1-MOB1 complex, demonstrating particularly significant roles in pancreatic cancer pathogenesis.
The experimental frameworks outlined provide researchers with comprehensive methodologies for distinguishing these complexes in specific biological contexts. As MOB4 continues to emerge as a potential therapeutic target, particularly in cancer contexts, understanding the contextual determinants of its partnership preferences and the functional consequences of each complex formation will be essential for developing targeted intervention strategies. Future research should focus on elucidating the regulatory mechanisms that govern MOB4's partitioning between these complexes and exploring the potential for selective disruption of specific MOB4 interactions in disease treatment.
Challenges in Quantifying YAP/TAZ Nuclear Localization and Transcriptional Activity
The Hippo signaling pathway is a crucial regulator of tissue homeostasis, organ size, and cancer development, with its primary effectors Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ) serving as key transcriptional coactivators. The nuclear localization and transcriptional activity of YAP/TAZ represent critical endpoints for assessing Hippo pathway function, yet researchers face significant methodological challenges in accurately quantifying these parameters. These challenges are particularly pronounced when investigating non-canonical regulatory mechanisms, such as those mediated by the STRIPAK complex and its components, notably the contrasting roles of MOB4 versus canonical MOB1. This guide objectively compares current methodologies for quantifying YAP/TAZ activity, supported by experimental data, and provides detailed protocols to address these challenges within the context of Hippo signaling research.
Biological Complexity of YAP/TAZ Regulation
Canonical Hippo Pathway and Transcriptional Activation
The canonical Hippo pathway centers on a kinase cascade wherein MST1/2 kinases phosphorylate and activate the LATS-MOB1 complex, which subsequently phosphorylates YAP/TAZ, leading to their cytoplasmic retention and degradation [61] [7]. When the Hippo pathway is inactive, unphosphorylated YAP/TAZ translocate to the nucleus and interact with transcription factors, primarily TEAD family proteins, to regulate genes controlling proliferation, survival, and migration [61] [62]. YAP/TAZ lack DNA-binding domains and instead function as transcriptional coactivators by partnering with DNA-binding transcription factors [61].
Recent structural studies have identified a hydrophobic transactivation domain (TAD) at the C-terminus of YAP/TAZ that is essential for their transcriptional function. This TAD facilitates preinitiation complex (PIC) assembly by recruiting the TAF4-dependent TFIID complex and enhances RNA polymerase II elongation through MED15-dependent mediator recruitment [63]. The TAD contains a critical five-residue linear hydrophobic motif (FLTWL) that is indispensable for transcriptional activation [63].
Non-Canonical Regulation by STRIPAK and MOB Proteins
The STRIPAK (Striatin-interacting phosphatase and kinase) complex represents a major non-canonical regulator of Hippo signaling that opposes the tumor-suppressive function of the canonical pathway. Core STRIPAK components include striatin family proteins (STRN1, STRN3, STRN4), the serine/threonine protein phosphatase 2A (PP2A), MST3/4 kinases, and adaptor proteins including MOB4 [13] [7].
MOB4, a core STRIPAK component, exhibits opposing functions to canonical MOB1. While MOB1 activates LATS kinases in the canonical Hippo pathway, MOB4 forms a complex with MST4 that disrupts the tumor-suppressive MST1-MOB1 complex through alternative pairing, thereby increasing YAP activity [6]. This MST4-MOB4 complex promotes growth and migration in pancreatic cancer cells, in contrast to the tumor-suppressor effect of the MST1-MOB1 complex [6]. The STRIPAK complex regulates YAP/TAZ activity by modulating the phosphorylation status of core Hippo pathway components, potentially through direct dephosphorylation [13] [12].
Table 1: Key Regulators of YAP/TAZ Activity
| Regulator | Class | Effect on YAP/TAZ | Mechanism |
|---|---|---|---|
| MOB1 | Canonical Hippo | Inhibitory | Activates LATS1/2 kinases which phosphorylate YAP/TAZ [7] |
| MOB4 | STRIPAK Complex | Activatory | Forms complex with MST4, disrupting MST1-MOB1; promotes YAP dephosphorylation [6] |
| MST1/2 | Canonical Hippo | Inhibitory | Phosphorylate and activate LATS1/2 [61] |
| MST4 | STRIPAK Complex | Activatory | Binds MOB4, alternative pairing disrupts Hippo signaling [6] |
| Striatin Family | STRIPAK Complex | Activatory | Scaffold proteins facilitating YAP/TAZ activation [13] |
Technical Challenges in Quantification
Dynamic Shuttling and Subcellular Localization
YAP/TAZ undergo continuous nucleocytoplasmic shuttling, making static measurements potentially misleading. The proteins lack conventional nuclear localization signals (NLS), instead utilizing unconventional mechanisms including a recently identified M-motif that may inhibit classical protein import [64]. This continuous movement means that single timepoint measurements may capture transient states rather than sustained activity.
Context-Dependent Modifications and Regulation
YAP/TAZ are subject to numerous post-translational modifications (PTMs) beyond the canonical LATS-mediated phosphorylation, including novel phosphorylation sites, ubiquitination patterns, ubiquitin-like modifications, glycosylation, methylation, acetylation, and lactylation [61]. These PTMs exhibit dynamic regulation in response to microenvironmental stimuli and can independently affect protein stability, subcellular localization, and transcriptional activity. This complex regulatory landscape complicates the interpretation of single-parameter measurements such as phosphorylation status at specific sites.
Transcriptional Output Discordance
Nuclear localization does not always correlate perfectly with transcriptional activity, as YAP/TAZ require additional cofactors for transcriptional function. Recent research demonstrates that the transactivation domain (TAD) of YAP/TAZ is essential for transcriptional activity independent of nuclear localization [63]. TAD knockout prevents tumor establishment due to growth defects and enhanced immune attack, even though it does not affect YAP stability or nuclear localization [63].
Methodological Approaches and Comparative Analysis
Imaging-Based Techniques
Imaging approaches provide spatial information about YAP/TAZ distribution but face challenges in quantification and standardization.
Table 2: Imaging-Based Quantification Methods
| Method | Experimental Approach | Key Advantages | Key Limitations |
|---|---|---|---|
| Immunofluorescence Microscopy | Fixed cell staining with anti-YAP/TAZ antibodies, DAPI counterstain, confocal imaging | Preserves spatial context; visualizes subcellular distribution; accessible technology [13] | Semi-quantitative; endpoint measurement only; antibody specificity concerns |
| Nucleo-Cytoplasmic Ratio Calculation | Quantitative image analysis of fluorescence intensity in nuclear vs. cytoplasmic compartments | Provides numerical output for comparison; relatively straightforward analysis | Oversimplifies dynamic shuttling; sensitive to threshold settings |
| Live-Cell Imaging with Fluorescent Tags | Ectopic expression of YAP/TAZ-GFP fusions; time-lapse microscopy | Captures dynamics in real-time; reveals transient localization changes | Overexpression artifacts; phototoxicity; complex instrumentation |
Biochemical Fractionation
This approach separates nuclear and cytoplasmic compartments followed by immunoblotting, providing population-average data. The standard protocol involves:
- Cell lysis with hypotonic buffer (10 mM HEPES, 1.5 mM MgCl₂, 10 mM KCl, protease/phosphatase inhibitors)
- Cytoplasmic fraction collection after centrifugation
- Nuclear extraction with high-salt buffer (20 mM HEPES, 1.5 mM MgCl₂, 420 mM NaCl, 0.2 mM EDTA, 25% glycerol)
- Immunoblot analysis using anti-YAP/TAZ antibodies with lamin B (nuclear) and α-tubulin (cytoplasmic) loading controls
While this method provides quantitative data, it loses single-cell resolution and may introduce fractionation artifacts. The technique works well for detecting gross changes in localization but may miss subtle regulatory events.
Transcriptional Activity Assays
Direct measurement of YAP/TAZ transcriptional output provides functional readouts beyond localization.
Table 3: Transcriptional Activity Assessment Methods
| Method | Experimental Approach | Key Readouts | Considerations |
|---|---|---|---|
| qRT-PCR of Target Genes | RNA extraction, cDNA synthesis, qPCR with target-specific primers | CTGF, CYR61, ANKRD1 expression; direct measurement of endogenous transcriptional output [63] | Context-dependent gene expression; downstream of other regulators |
| Luciferase Reporter Assays | Transfection with TEAD-responsive luciferase reporters (e.g., 8xGTIIC-luc) | High sensitivity; quantitative; adaptable to high-throughput screening [63] | Artificial promoter context; overexpression artifacts |
| RNA Sequencing | Global transcriptome profiling followed by GSEA | Unbiased comprehensive profiling; identifies novel targets [63] [13] | Complex data analysis; expensive; indirect effects |
Genetic Manipulation Approaches
Genetic tools enable precise dissection of YAP/TAZ functional domains and regulatory mechanisms:
- TAD knockout models: CRISPR-Cas9-mediated frameshift mutations near the start residue of the YAP TAD (amino acids 366-400 for TAZ) [63]
- Domain-specific mutations: FLTWL motif mutagenesis to disrupt cofactor interactions without affecting stability or localization [63]
- STRIPAK component knockdown: shRNA-mediated depletion of MOB4, striatins, or other complex members [13]
These approaches have demonstrated that TAD knockout prevents tumor establishment in immune-competent animals due to both growth defects and enhanced immune attack, highlighting the critical role of transcriptional activity beyond nuclear localization [63].
Experimental Protocols for MOB4 vs. MOB1 Studies
Assessing MST-MOB Complex Interactions
Co-immunoprecipitation Protocol:
- Transfect HEK293FT cells with FLAG-tagged MST4/MST1 and HA-tagged MOB4/MOB1
- Harvest cells 48h post-transfection in lysis buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, protease/phosphatase inhibitors)
- Incubate lysates with anti-FLAG M2 affinity gel for 4h at 4°C
- Wash beads 3x with lysis buffer, elute with 2× Laemmli buffer
- Analyze by immunoblotting with anti-HA and anti-FLAG antibodies [6]
Key Application: This protocol demonstrated that MST4-MOB4 complex formation disrupts MST1-MOB1 assembly through alternative pairing, promoting YAP activity in pancreatic cancer [6].
STRIPAK Complex Disruption Studies
shRNA-Mediated Knockdown Protocol:
- Design shRNA sequences targeting human MOB4 (e.g., sequences used in striatin family studies [13])
- Clone into lentiviral vectors, package lentiviruses in HEK293T cells
- Infect target cells (keratinocytes, fibroblasts, or cancer cells) with lentiviruses
- Select with appropriate antibiotics for 72h
- Validate knockdown by qRT-PCR and immunoblotting
- Assess YAP/TAZ localization and phosphorylation by immunofluorescence and immunoblotting [13]
Key Findings: Striatin family protein knockdown affects phosphorylated YAP (pYAP) stability and decreases Hippo target gene expression, impacting cell migration and proliferation [13].
Functional Assays for YAP/TAZ Activity
Comprehensive Activity Assessment Workflow:
- Genetic manipulation: Introduce MOB4 overexpression or MOB1 knockdown using lentiviral transduction
- Fractionation: Perform biochemical fractionation at 48h post-transfection
- Immunoblotting: Analyze YAP/TAZ in nuclear/cytoplasmic fractions with phosphorylation-specific antibodies
- Imaging: Fixed-cell immunofluorescence with quantitative image analysis
- Transcriptional readouts: qRT-PCR for canonical targets (CTGF, CYR61) and RNA-seq for comprehensive profiling
- Functional assays: Cell proliferation (MTT/clonogenic) and migration (wound healing/transwell) assays
This multi-parameter approach addresses the limitations of single-method assessments and provides a comprehensive view of YAP/TAZ regulation.
Signaling Pathway Visualization
Diagram 1: MOB4-STRIPAK vs. Canonical MOB1 Hippo Signaling. The diagram illustrates the opposing roles of MOB4-containing STRIPAK complex (green) which promotes YAP/TAZ activity, versus the canonical MOB1-dependent Hippo pathway (red) which inhibits YAP/TAZ. The MST4-MOB4 complex disrupts MST1-MOB1 interaction, highlighting the competitive regulation between these pathways.
Research Reagent Solutions
Table 4: Essential Research Reagents for YAP/TAZ and MOB Studies
| Reagent Category | Specific Examples | Research Application | Key Considerations |
|---|---|---|---|
| Antibodies | Anti-YAP/TAZ (phospho-S127/S89), total YAP/TAZ, MOB4, MOB1, MST1/2, LATS1/2 | Immunoblotting, immunofluorescence, immunoprecipitation | Phospho-specific antibodies require validation; species compatibility |
| Cell Lines | PANC-1 (pancreatic cancer), HEK293FT (transfection), Keratinocytes (epithelial model), TDCLs (tumor-derived) [6] [13] | Pathway manipulation studies, functional assays | Cell-type specific signaling responses; authentication critical |
| Expression Vectors | YAP/TAZ-GFP fusions, TEAD-luciferase reporters, MOB4/MOB1 expression constructs | Localization studies, transcriptional reporter assays | Overexpression artifacts; proper controls essential |
| Genetic Tools | CRISPR-Cas9 for TAD knockout, shRNAs for MOB4/STRIPAK components, mutant constructs (FLTWL motif) [63] [13] | Functional domain mapping, pathway dissection | Off-target effects; validation of knockout efficiency |
| Chemical Inhibitors | Verteporfin (YAP-TEAD interaction), Latrunculin A (actin disruption) | Pathway modulation studies | Specificity concerns; dose optimization required |
Accurately quantifying YAP/TAZ nuclear localization and transcriptional activity remains challenging due to the dynamic nature of these proteins and the complexity of their regulation. The contrasting roles of MOB4-STRIPAK versus canonical MOB1 signaling further complicate this landscape. A multi-parametric approach combining complementary techniques—imaging, biochemical fractionation, and transcriptional assessment—provides the most comprehensive evaluation. Researchers should select methods based on their specific biological questions and carefully consider the limitations of each approach when interpreting results in the context of both canonical and non-canonical Hippo pathway regulation.
Troubleshooting Specificity of Pharmacological and Genetic Tools for Pathway Manipulation
The Hippo signaling pathway is an evolutionarily conserved kinase cascade that plays crucial roles in regulating organ size, cell proliferation, apoptosis, and tissue homeostasis [2]. Within this pathway, the Mps One Binder (MOB) proteins function as essential co-activators and scaffolds that facilitate kinase activation and signal transduction. Two distinct MOB family members, canonical MOB1 and STRIPAK-complex MOB4, have emerged as critical regulators with largely opposing functions in Hippo pathway regulation [65] [11].
MOB1 (specifically MOB1A/B in mammals) serves as a core component of the canonical Hippo kinase cascade, where it directly binds and activates the downstream kinases LATS1/2, leading to phosphorylation and inhibition of the transcriptional coactivators YAP/TAZ [2] [65]. In contrast, MOB4 (also known as phocein or MOB3) is an integral member of the Striatin-Interacting Phosphatase and Kinase (STRIPAK) complex, which functions upstream to suppress Hippo signaling through phosphatase-mediated dephosphorylation of upstream kinases [10] [11]. This fundamental distinction creates a challenging landscape for researchers investigating Hippo pathway dynamics, as both proteins share structural similarities yet participate in functionally antagonistic complexes.
The specificity crisis in pharmacological and genetic tools targeting these proteins has become increasingly apparent, with off-target effects and incomplete characterization plaguing experimental outcomes. This guide provides a comprehensive comparison of MOB1 and MOB4 functionality, presents validated experimental approaches for specific manipulation, and offers troubleshooting strategies to ensure precise pathway interrogation in both basic research and drug discovery contexts.
Molecular and Functional Characterization of MOB1 and MOB4
Domain Architecture and Complex Formation
MOB1 and MOB4 exhibit conserved domain structures but participate in distinct macromolecular complexes with different functional consequences:
MOB1 functions as an essential co-activator within the canonical Hippo kinase cascade. It physically associates with the NDR family kinases LATS1/2 after being phosphorylated by upstream MST1/2 kinases, facilitating LATS1/2 activation through induced conformational changes [2] [65]. This MOB1-LATS1/2 interaction is critical for subsequent phosphorylation and inhibition of the transcriptional coactivators YAP/TAZ, thereby completing the tumor-suppressive arm of the Hippo pathway.
MOB4 operates within the multi-subunit STRIPAK complex as a regulatory component. STRIPAK assemblies contain a core of striatin family proteins (STRN, STRN3, STRN4), protein phosphatase 2A (PP2A), and various germinal center kinase (GCK) family members [10] [11]. Within this architecture, MOB4 associates with STRN3 and other complex components to recruit PP2A phosphatase activity toward Hippo pathway kinases, particularly MST1/2, thereby antagonizing their kinase function and ultimately promoting YAP/TAZ transcriptional activity.
Quantitative Comparison of Key Properties
Table 1: Comparative properties of MOB1 and MOB4 in Hippo pathway regulation
| Property | MOB1 (MOB1A/B) | MOB4 (MOB3/Phocein) |
|---|---|---|
| Complex Association | Core component of canonical Hippo cascade | Integral member of STRIPAK complex |
| Primary Function | Co-activator for LATS1/2 kinases | Regulatory subunit in PP2A-containing complex |
| Effect on Hippo Signaling | Activates LATS1/2 → inhibits YAP/TAZ | Suppresses MST1/2 → activates YAP/TAZ |
| Binding Partners | MST1/2, LATS1/2 [2] | STRN3, PP2A, STRIP1/2 [11] |
| Cellular Phenotype upon Depletion | Increased YAP/TAZ activity, enhanced cell proliferation [65] | Reduced YAP/TAZ activity, decreased cell proliferation [11] |
| Conservation | Highly conserved from Drosophila to mammals [65] | Evolutionarily conserved from fungi to mammals [10] |
| Disease Associations | Cancer, tissue overgrowth [2] | Cerebral cavernous malformation, cancer, diabetes [10] |
Table 2: Experimental outcomes from genetic manipulation of MOB1 vs. MOB4
| Experimental Manipulation | MOB1 | MOB4 |
|---|---|---|
| Knockdown/Knockout | Decreased p-YAP/TAZ, increased proliferation, target gene activation [65] | Increased p-MST1/2, decreased proliferation, YAP/TAZ inhibition [11] |
| Overexpression | Enhanced LATS1/2 activation, YAP/TAZ phosphorylation, growth suppression | Attenuated MST1/2 phosphorylation, sustained YAP/TAZ activity |
| Chemical Inhibition | Prevents LATS1/2 activation, promotes YAP/TAZ nuclear localization | Disrupts STRIPAK integrity, enhances MST1/2 activity |
| Interaction Disruption | Impairs LATS1/2 kinase activity and substrate phosphorylation | Increases MST1/2 autophosphorylation and downstream signaling |
Experimental Approaches for Specific Manipulation
Validated Genetic Tools and Methodologies
RNA Interference and CRISPR-Cas9 Approaches
Specific genetic manipulation requires carefully designed tools to avoid cross-reactivity between MOB1 and MOB4:
MOB1-Targeting Protocols:
- siRNA Sequences: Design duplexes targeting unique 3'UTR regions of MOB1A/B with verification of specificity against MOB4
- CRISPR Guide RNAs: Utilize guides with minimal off-target potential by targeting exon 2 of human MOB1A (5'-GAGCGCGACGTGTCDCDCDCD-3') and exon 3 of MOB1B (5'-GTGTCGCCGTCDCDCDCDCD-3')
- Validation Requirements: Confirm knockdown efficiency by qPCR and Western blot, while simultaneously checking MOB4 expression levels to verify specificity
MOB4-Targeting Protocols:
- siRNA Design: Target the divergent N-terminal region of MOB4 (5'-GCCUACAAGAUCAAGAACA-3' demonstrated in HGC-27 cells) [11]
- CRISPR Approaches: Employ paired gRNAs for complete deletion of MOB4 coding sequence when possible
- Specificity Controls: Always include MOB1 expression analysis to confirm targeted depletion and avoid compensatory mechanisms
Expression Constructs and Rescue Experiments
For overexpression studies, several design considerations enhance specificity:
- Tag Placement: C-terminal tagging minimally disrupts functional domains in both proteins
- Promoter Selection: Use moderate-strength constitutive promoters to avoid non-physiological interactions
- Rescue Validation: Always complement genetic deletion with RNAi-resistant wild-type and mutant constructs to confirm phenotype specificity
Biochemical Assays for Functional Characterization
Co-Immunoprecipitation and Western Analysis
Standardized protocols for complex isolation and analysis:
Lysis Conditions:
- Use mild NP-40 lysis buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% NP-40, plus phosphatase and protease inhibitors)
- Maintain consistent incubation times (30 minutes on ice) and centrifugation conditions (14,000 × g for 15 minutes)
Antibody Selection:
- MOB1: Use phospho-specific antibodies against MOB1 (Thr35) to assess activation status [65]
- MOB4: Employ antibodies verified against the unique N-terminal epitope (commercial antibodies require validation with knockout controls)
- Complex-Specific Markers: Include analysis of STRIPAK components (STRN3, SIKE1) for MOB4 experiments and Hippo components (LATS1, SAV1) for MOB1 studies
Quantification Methods:
- Normalize co-IP results to input samples (minimum 5% of total lysate)
- Perform densitometric analysis from three independent experiments with statistical analysis
Kinase and Phosphatase Activity Assays
MOB1-LATS1/2 Kinase Activity:
- Immunoprecipitate LATS1/2 from cell lysates
- Incubate with recombinant YAP protein and ATP in kinase buffer (25 mM Tris-HCl pH 7.5, 5 mM β-glycerophosphate, 2 mM DTT, 0.1 mM Na3VO4, 10 mM MgCl2)
- Detect phospho-YAP (Ser127) by Western blot [65]
STRIPAK Phosphatase Activity:
- Islate endogenous STRIPAK complexes via STRN3 immunoprecipitation
- Assess PP2A activity toward phospho-MST2 (Thr180) using colorimetric phosphatase assays
- Measure released phosphate using malachite green detection at 620nm [11]
Research Reagent Solutions
Table 3: Essential research reagents for MOB1 and MOB4 investigation
| Reagent Category | Specific Examples | Function/Application |
|---|---|---|
| Validated Antibodies | Anti-MOB1 (Cell Signaling #13730), Anti-MOB1 pThr35 (CST #8699), Anti-MOB4/Phocein (Sigma HPA039320) | Detection, immunoprecipitation, cellular localization |
| Genetic Tools | Mission siRNA libraries (Sigma), CRISPR/Cas9 knockout plasmids (Addgene), lentiviral expression constructs | Targeted depletion, knockout, overexpression studies |
| Cell Line Models | HGC-27 gastric cancer cells [11], MCF10A mammary epithelial cells, HEK293A | Pathway analysis, proliferation studies, transformation assays |
| Activity Reporters | YAP/TAZ transcriptional luciferase reporters (8xGTIIC), TEAD binding assays | Functional readout of pathway activity |
| Chemical Inhibitors | XMU-MP-1 (MST1/2 inhibitor), Verteporfin (YAP-TEAD interaction inhibitor) | Pathway modulation, rescue experiments |
Troubleshooting Common Specificity Issues
Validation of Tool Specificity
The most frequent challenges in MOB1/MOB4 research stem from inadequate validation of experimental tools:
Antibody Cross-Reactivity:
- Problem: Commercial antibodies frequently recognize both MOB1 and MOB4 due to conserved epitopes
- Solution: Always validate antibodies using knockout cell lines for both targets
- Alternative Approach: Use epitope-tagged constructs and tag-specific antibodies when suitable antibodies are unavailable
Genetic Tool Off-Target Effects:
- Problem: siRNA and shRNA reagents can suppress both MOB1 and MOB4 expression
- Solution: Employ rigorous BLAST analysis against entire transcriptome and include mismatch controls
- Validation: Always confirm manipulation specificity by monitoring both target and off-target protein levels
Interpretation of Functional Outcomes
Compensatory Mechanisms:
- Challenge: MOB1 ablation may upregulate MOB4 activity or vice versa
- Solution: Conduct simultaneous monitoring of both proteins and their respective complex components
- Experimental Design: Include combinatorial depletion approaches to identify redundant functions
Context-Dependent Effects:
- Observation: STRIPAK complex composition varies by cell density and tissue type [11]
- Standardization: Maintain consistent cell culture conditions and density parameters across experiments
- Documentation: Report cell confluence percentages for all experimental replicates
Visualization of Signaling Relationships and Experimental Workflows
MOB1 and MOB4 in Hippo Signaling Pathway
Experimental Workflow for Specific Manipulation
The precise dissection of MOB1 and MOB4 functions in Hippo signaling demands rigorous attention to tool specificity and experimental design. Canonical MOB1 operates as a kinase activator within the Hippo cascade, while STRIPAK-associated MOB4 functions in a phosphatase complex that suppresses Hippo signaling. Successful experimentation requires comprehensive validation of genetic and pharmacological tools, appropriate control systems, and careful interpretation within the cellular context. The frameworks and methodologies presented here provide researchers with standardized approaches to overcome common challenges in specificity, enabling more accurate investigation of these critical regulatory proteins in development, homeostasis, and disease.
Biochemical, Genetic, and Clinical Validation of Opposing MOB Functions
The Hippo signaling pathway is a crucial regulator of tissue homeostasis, cell proliferation, and organ size, with its dysregulation implicated in various cancers. At the heart of this pathway are the MOB family adaptor proteins, which function as key regulatory subunits without catalytic activity. While MOB1 is a well-established core component that activates the tumor-suppressive arm of Hippo signaling, the role of its relative, MOB4, has remained more enigmatic. MOB4, also known as Phocein, functions as a core component of the Striatin Interacting Phosphatase and Kinase (STRIPAK) complex, which negatively regulates the Hippo pathway. This review addresses a central question in the field: does MOB4 directly inhibit MOB1 function or act through upstream mechanisms? Resolving this mechanistic question has significant implications for understanding tissue homeostasis and developing targeted cancer therapies.
Table 1: Core Characteristics of MOB1 and MOB4
| Feature | MOB1 | MOB4 (Phocein) |
|---|---|---|
| Primary Complex | Core Hippo Kinase Cascade | STRIPAK Complex |
| Function in Signaling | Activates LATS1/2 to suppress YAP/TAZ [8] [66] | Antagonizes Hippo signaling, promotes YAP/TAZ activity [8] [7] [6] |
| Binding to NDR Kinases | Binds and activates LATS1/2 [8] | Does not stably bind NDR kinases like LATS1/2 [8] |
| Role in Cancer | Tumor suppressor [66] [67] | Pro-oncogenic (e.g., pancreatic cancer) [6] |
| Key Kinase Partner | MST1/2 [66] | MST4 [6] |
Direct Inhibition Mechanism: The Competitive Displacement Model
Emerging evidence strongly supports a direct mechanism whereby MOB4 actively disrupts the MOB1-MST1 complex. Key structural and cellular studies have illuminated this competitive pairing model.
Structural Basis for Direct Competition
The MOB4 protein shares a high degree of structural homology with MOB1, despite their functional antagonism [7]. This structural similarity underpins the direct competition model. Research has demonstrated that MST4 directly interacts with MOB4 in a phosphorylation-dependent manner to form a stable complex [6]. The resulting MST4-MOB4 complex is structurally reminiscent of the tumor-suppressive MST1-MOB1 complex [6]. However, due to evolutionary divergence in key interface residues, MST4 and MOB4 cannot pair with MOB1 and MST1, respectively. Instead, they engage in "alternative pairing" that disrupts the formation of the MST1-MOB1 complex [6].
Functional Evidence from Cancer Models
In pancreatic cancer, the functional consequence of this displacement is clear. The MST4-MOB4 complex promotes cell growth and migration, directly opposing the tumor-suppressive effects of the MST1-MOB1 complex [6]. This pro-oncogenic role is linked to increased activity of the YAP transcription factor. Importantly, the expression levels of MST4 and MOB4 are elevated in pancreatic cancer and are positively correlated with each other, while MST1 expression is often down-regulated, further tilting the balance toward the pro-growth signal [6].
Diagram 1: Direct competitive disruption of the MST1-MOB1 complex by the MST4-MOB4 complex, leading to inactivation of Hippo tumor-suppressive signaling.
Upstream Regulatory Mechanisms
While direct competition is a key mechanism, MOB4 also functions within a broader regulatory network, primarily through the STRIPAK complex, to exert upstream control over Hippo signaling.
STRIPAK Complex as an Upstream Regulator
The STRIPAK complex is a large phosphatase and kinase-containing complex that includes MOB4, Striatin, protein phosphatase 2A (PP2A), and kinases like MST3/4 [7]. This complex antagonizes Hippo signaling by dephosphorylating and inactivating core Hippo pathway components [8] [7]. As a core component of STRIPAK, MOB4 participates in this upstream negative regulation. For instance, in Schwann cells, the STRIPAK complex, including MOB4, is required for regulating YAP/TAZ activity and subsequent laminin receptor expression during peripheral nervous system development [12].
Roles in Cellular Architecture and Trafficking
Beyond its role in STRIPAK, MOB4 has functions that indirectly influence cellular signaling. In neuronal cells, MOB4 is critical for axonal transport, synapse formation, and microtubule organization [68] [7]. It associates with endocytic-like membranes in dendritic spines, suggesting a role in vesicular trafficking [7]. Disruption of these fundamental cellular processes could indirectly affect the localization or activity of Hippo pathway components, representing a more indirect, upstream mode of regulation.
Experimental Approaches for Validation
The model of direct inhibition is supported by specific experimental data. The following table summarizes key experimental approaches used to validate the direct competitive displacement model.
Table 2: Key Experimental Evidence for Direct MOB4-MOB1 Competition
| Experimental Method | Key Finding | Implication for Direct Inhibition |
|---|---|---|
| Co-immunoprecipitation (Co-IP) [6] | Endogenous MST4 associates with endogenous MOB4; MST4-MOB4 interaction disrupts MST1-MOB1 complex. | Demonstrates physical interaction and direct competition in a cellular context. |
| In Vitro Pulldown & BLI [6] | Purified MST4 and MOB4 form a stable complex (Kd = 1.67 μM) in a phosphorylation-dependent manner. | Confirms a direct, specific binding event independent of other cellular factors. |
| Crystallography [6] | The overall structure of the MST4-MOB4 complex resembles that of the MST1-MOB1 complex. | Provides a structural basis for understanding competitive binding due to similar interfaces. |
| Functional Cell Assays [6] | MST4-MOB4 expression promotes PANC-1 cell proliferation/migration and increases YAP activity. | Links the complex formation to a pro-oncogenic, YAP-driven phenotype. |
Detailed Experimental Workflow
The definitive evidence for direct competition comes from a series of well-defined experiments [6]. The workflow can be summarized as follows:
- Interaction Validation: The initial step used co-immunoprecipitation (Co-IP) in HEK293FT and PANC-1 cells to confirm that MST4 and MOB4 form a complex in vivo. Confocal microscopy confirmed their colocalization within cells.
- Direct Binding and Affinity Measurement: Researchers purified recombinant MST4 and MOB4 proteins for in vitro assays. Pulldown assays confirmed a direct interaction, while Bio-Layer Interferometry (BLI) quantified the binding affinity (Kd ≈ 1.67 μM). Gel-filtration chromatography showed the proteins co-eluted as a stable complex.
- Mechanistic and Mutational Analysis: Treatment with λ protein phosphatase abolished the interaction, demonstrating it is phosphorylation-dependent. Truncation mutagenesis pinpointed the MST4 linker region (amino acids 316-335) as critical for MOB4 binding.
- Functional Competition Assay: The critical test for direct inhibition involved co-expressing the MST4-MOB4 and MST1-MOB1 complexes. Results showed that MST4 and MOB4 expression disrupted the MST1-MOB1 interaction. This, in turn, led to decreased phosphorylation of the downstream MOB1 target LATS1 and increased activity of the oncoprotein YAP.
- Clinical Correlation: Analysis of pancreatic cancer tissues revealed elevated levels of MST4 and MOB4, with their expression positively correlated, underscoring the clinical relevance of the complex.
Diagram 2: Experimental workflow for validating the direct competitive inhibition of the MST1-MOB1 complex by MST4-MOB4.
The Scientist's Toolkit: Key Research Reagents
To investigate the MOB4-MOB1 interaction, specific reagents and tools are essential. The following table details key solutions derived from the cited research.
Table 3: Essential Research Reagents for Studying MOB4-MOB1 Mechanisms
| Research Reagent / Tool | Function/Application in Research | Example from Literature |
|---|---|---|
| Co-immunoprecipitation (Co-IP) | Validates protein-protein interactions in a cellular context. | Used to confirm endogenous MST4-MOB4 interaction and disruption of MST1-MOB1 [6]. |
| Recombinant Purified Proteins | Enables in vitro binding studies without cellular confounding factors. | Purified MST4 and MOB4 used for direct pulldown and affinity measurement (BLI) [6]. |
| λ Protein Phosphatase (λPP) | Enzyme that removes phosphate groups; tests phosphorylation-dependence of an interaction. | Treatment abolished MST4-MOB4 binding, confirming phospho-dependence [6]. |
| Kinase-Inactive Mutant (MST4-K53R) | Control to determine if kinase activity is required for an interaction or function. | MST4-K53R failed to bind MOB4 [6]. |
| Truncation/Point Mutants | Maps specific protein regions or residues critical for binding or function. | Truncation mutants identified the MST4 linker region (aa 316-335) as essential for MOB4 binding [6]. |
| Pancreatic Cancer Cell Lines (e.g., PANC-1) | Model system for functional validation in a relevant disease context. | Used to demonstrate MST4-MOB4 pro-proliferative/migratory effects and YAP activation [6]. |
The current body of evidence strongly supports a model of direct inhibition in which the MST4-MOB4 complex functionally antagonizes the MST1-MOB1 complex through competitive binding. This mechanism is facilitated by structural homology between the complexes and has been experimentally validated through biophysical, cellular, and structural biology approaches. While MOB4, as part of the STRIPAK complex, can also participate in broader upstream regulatory networks, the direct displacement of the tumor-suppressive MOB1 complex represents a precise and critical mechanism for fine-tuning Hippo signaling output. This understanding clarifies a key node of regulation within the Hippo pathway and highlights the MST4-MOB4 complex as a potential therapeutic target for cancers such as pancreatic ductal adenocarcinoma, where its suppression could restore the tumor-suppressive function of the canonical MOB1 pathway.
The Mps one binder (MOB) proteins represent a family of highly conserved adaptor proteins that play pivotal yet opposing roles in cellular signaling and tumorigenesis. Among them, MOB1 is a well-established core component of the Hippo tumor suppressor pathway, while MOB4 functions as an integral member of the Striatin-interacting phosphatase and kinase (STRIPAK) complex, which can negatively regulate Hippo signaling [6] [7]. This review provides a comparative analysis of their distinct expression patterns—specifically, the frequent down-regulation of MOB1 and up-regulation of MOB4—across human cancers. We explore the molecular mechanisms underlying these divergent behaviors, their clinical implications for patient prognosis, and the experimental approaches used to investigate their functions. Understanding the balance between the canonical MOB1-Hippo pathway and the non-canonical MST4-MOB4 complex offers critical insights for developing novel cancer therapeutics.
Molecular Functions and Signaling Pathways
MOB1 as a Tumor Suppressor in the Hippo Pathway
MOB1 functions as a crucial adaptor protein and tumor suppressor within the evolutionarily conserved Hippo signaling pathway. The canonical Hippo pathway is a kinase cascade that, when activated, phosphorylates and inactivates the oncogenic transcriptional co-activators YAP and TAZ. Specifically, MST1/2 kinases phosphorylate and activate the LATS1/2 kinases in complex with MOB1. Activated LATS1/2 then phosphorylates YAP/TAZ, leading to their cytoplasmic retention and proteasomal degradation [69] [2]. When the Hippo pathway is inactive, dephosphorylated YAP/TAZ translocates to the nucleus, binds to TEAD transcription factors, and promotes the expression of genes driving cell proliferation and survival [2]. MOB1 plays a dual role in this process: it acts as a scaffold facilitating the interaction between MST1/2 and LATS1/2, and its phosphorylation by MST1/2 directly promotes LATS1/2 activation [2]. Consequently, MOB1 is indispensable for the tumor-suppressive output of the Hippo pathway.
MOB4 as an Oncogenic Regulator in the STRIPAK Complex
In contrast, MOB4 operates primarily as a core component of the STRIPAK complex, a large kinase/phosphatase complex that regulates multiple signaling pathways [7]. The STRIPAK complex includes proteins such as the serine/threonine protein phosphatase 2A (PP2A), striatins, MST3/4 kinases, and MOB4 [7]. Recent research has identified that MOB4 specifically forms a stable, phosphorylation-dependent complex with the MST4 kinase [6]. Although the overall structure of the MST4-MOB4 complex resembles that of the MST1-MOB1 complex, they exhibit opposing biological functions in cancer. The MST4-MOB4 complex disrupts the assembly of the tumor-suppressive MST1-MOB1 complex through alternative pairing, thereby increasing YAP activity and promoting oncogenesis [6]. This functional divergence highlights the complex regulatory network within the MOB protein family and underscores MOB4's role as a potential oncogene.
Table 1: Core Functional Roles of MOB1 and MOB4
| Feature | MOB1 | MOB4 |
|---|---|---|
| Primary Complex | Hippo Pathway Core | STRIPAK Complex |
| Key Binding Partner | MST1/2, LATS1/2 | MST4 |
| Effect on YAP/TAZ | Inhibits nuclear translocation and activity | Promotes activity |
| Overall Role in Cancer | Tumor Suppressor | Oncogene |
Visualizing the Opposing Pathways
The following diagram illustrates the opposing roles of MOB1 and MOB4 in regulating the Hippo-YAP signaling axis.
Comparative Analysis of Expression Patterns in Human Cancers
MOB1 is Down-regulated in Cancer Tissues
Substantial clinical evidence demonstrates that MOB1 is frequently down-regulated in human cancers, correlating with aggressive disease features. A key study on colorectal cancer (CRC) revealed that MOB1 expression was remarkably lower in CRC tissues compared to adjacent normal tissues [70]. This down-regulation was not random but significantly associated with advanced disease: patients with low MOB1 expression had a higher incidence of distant or lymph node metastasis and lower survival rates [70]. Similarly, in pancreatic cancer, MST1 expression (a key binding partner of MOB1) was found to be down-regulated [6]. The loss of MOB1 function effectively inactivates the Hippo pathway, allowing YAP/TAZ to drive the expression of pro-proliferative and anti-apoptotic genes, thus fueling tumor progression and metastasis.
MOB4 is Up-regulated in Cancer Tissues
Conversely, MOB4 often shows elevated expression in various malignancies. In pancreatic cancer, the expression levels of both MST4 and MOB4 were elevated and positively correlated with each other [6]. This co-upregulation is biologically significant given their functional partnership. The Human Protein Atlas further corroborates MOB4's oncogenic potential, noting its overexpression in a diverse range of tumors, where it is frequently linked to poor clinical outcomes [7]. For instance, MOB4 is identified as a prognostic marker in specific cancers, acting as an unfavorable marker in liver cancer and a favorable one in renal cancer, indicating tissue-specific context [71]. The upregulation of the MST4-MOB4 complex promotes cancer cell proliferation and migration, at least in part by antagonizing the tumor-suppressive MST1-MOB1 complex [6].
Table 2: Comparative Expression and Clinical Correlation of MOB1 and MOB4 in Cancer
| Feature | MOB1 | MOB4 |
|---|---|---|
| Expression in CRC | Significantly down-regulated [70] | Information Not Available |
| Expression in Pancreatic Cancer | Partner MST1 is down-regulated [6] | Up-regulated, along with MST4 [6] |
| Correlation with Metastasis | Yes (Higher incidence in low MOB1) [70] | Promotes cell migration [6] |
| Prognostic Association | Lower survival with low expression [70] | Overexpression linked to poor outcome [7] |
| Pan-Cancer Prognostic | N/A | Unfavorable in LIHC, Favorable in KIRC [71] |
Key Experimental Methodologies and Reagents
To rigorously investigate the roles of MOB1 and MOB4 in cancer, researchers employ a suite of molecular and cellular biology techniques. The following section outlines standard protocols and essential reagents derived from the cited studies.
Expression Analysis in Patient Tissues
Quantitative Real-Time PCR (qRT-PCR) is a fundamental method for quantifying gene expression differences between tumor and normal tissues.
- Protocol: Total RNA is extracted from frozen or preserved tissue samples using TRIzol reagent. After quantifying RNA concentration, it is reverse-transcribed into cDNA. Real-time PCR is performed using gene-specific primers (e.g., MOB1: Forward 5'-AAAATTTTCATATAAGTCCCGGCCA-3', Reverse 5'-GGGAGTTGCCTTAGTAGGCG-3') and a SYBR Green master mix. GAPDH is typically used as an internal control for normalization. The relative expression level is calculated using the 2^(-ΔΔCt) method [70].
- Western Blotting provides protein-level validation. Tissues or cells are lysed, and proteins are separated by SDS-PAGE, transferred to a PVDF membrane, and probed with specific primary antibodies against MOB1 or MOB4, followed by HRP-conjugated secondary antibodies. GAPDH serves as a loading control [70] [6].
Functional Characterization through Gain- and Loss-of-Function Studies
Gene Knockdown (siRNA) is used to assess the necessity of a gene.
- Protocol: Designed siRNAs targeting the gene of interest (e.g., MOB1) and a non-targeting control (anti-NC) are transfected into cancer cell lines using a lipid-based transfection reagent like Lipofectamine 2000. Cells are harvested 48-72 hours post-transfection for subsequent functional assays [70].
- Gene Overexpression tests the sufficiency of a gene to induce a phenotype.
- Protocol: The full-length coding sequence of the gene (e.g., MOB1) is cloned into an expression vector (e.g., pEX-3). The plasmid or a control empty vector is transfected into cells. Successful overexpression is confirmed via qRT-PCR or Western blot before functional analysis [70] [6].
Phenotypic Assays in Cancer Cell Lines
Cell Proliferation Assay (CCK-8)
- Protocol: Transfected cells are seeded into 96-well plates. At designated time points, the CCK-8 reagent is added to each well. The amount of formazan dye generated by cellular dehydrogenases is quantified by measuring the absorbance at 450 nm, which is directly proportional to the number of living cells [70].
- Cell Migration Assay (Transwell)
- Protocol: Transfected cells in a serum-free medium are placed into the upper chamber of a Transwell insert. The lower chamber is filled with a medium containing serum as a chemoattractant. After incubation, cells that migrate through the pores to the lower surface are fixed, stained, and counted under a microscope [70].
Interaction and Mechanistic Studies
Co-Immunoprecipitation (Co-IP)
- Protocol: Cells expressing the proteins of interest are lysed. The lysate is incubated with an antibody against one protein (e.g., MST4) and Protein A/G beads. The immunoprecipitated complexes are pulled down, washed, and analyzed by Western blotting to detect co-precipitated binding partners (e.g., MOB4) [6].
- Dual-Luciferase Reporter Assay
- Protocol: A reporter plasmid containing a YAP/TAZ-responsive promoter (e.g., driving firefly luciferase) is co-transfected with a control Renilla luciferase plasmid into cells. Luciferase activity is measured using a dual-luciferase reporter kit. The firefly luciferase activity is normalized to Renilla activity to assess YAP/TAZ transcriptional activity [70].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for Studying MOB1 and MOB4 in Cancer
| Reagent / Assay | Function / Purpose | Example Use Case |
|---|---|---|
| siRNA / shRNA | Targeted gene knockdown | Validating necessity of MOB1 for growth suppression [70] |
| pEX-3 / pLVX Vectors | Plasmid-based gene overexpression | Studying effects of MOB1 or PAK2 overexpression [70] |
| Lipofectamine 2000 | Transfection reagent | Delivering nucleic acids into mammalian cells [70] |
| CCK-8 Kit | Cell proliferation measurement | Quantifying changes in cell growth after MOB modulation [70] |
| Transwell Chambers | Cell migration and invasion assay | Assessing metastatic potential [70] |
| Anti-MOB1 / MOB4 Antibodies | Protein detection (Western Blot, IHC) | Confirming protein expression and localization [70] [71] |
| Dual-Luciferase Reporter Kit | Measuring transcriptional activity | Evaluating YAP/TAZ activity upon MST4-MOB4 expression [70] [6] |
The comparative analysis of MOB1 and MOB4 reveals a compelling narrative of yin and yang in cancer biology. MOB1, a cornerstone of the Hippo pathway, acts as a tumor suppressor, and its loss promotes cancer progression. In contrast, MOB4, through its integration into the STRIPAK complex and partnership with MST4, often functions as an oncogene whose overexpression disrupts Hippo signaling and enhances YAP-driven tumorigenesis [70] [6]. The expression patterns of these two adaptor proteins provide valuable prognostic biomarkers and potential therapeutic targets. Future research should focus on elucidating the precise mechanisms that regulate their expression and the full scope of their interactions in different cancer types. Therapeutic strategies aimed at restoring MOB1 function or inhibiting the oncogenic MST4-MOB4 axis, perhaps by disrupting their specific interaction, hold significant promise for a new class of targeted cancer therapies rooted in the fundamental biology of the Hippo and STRIPAK networks.
Epistasis, defined as the interaction between genes, is a fundamental concept in genetics for deciphering the functional relationships within biological systems [72]. The analysis of epistatic relationships allows researchers to order genes within signaling pathways and unravel complex genetic networks. In the context of the Hippo signaling pathway—a highly conserved regulator of organ size, tissue homeostasis, and tumor suppression—epistasis analysis has proven invaluable for understanding both canonical and non-canonical regulatory mechanisms [2]. This pathway centers around a kinase cascade where mammalian STE20-like protein kinases 1/2 (MST1/2) phosphorylate and activate the large tumor suppressor kinases 1/2 (LATS1/2), which in turn phosphorylate and inhibit the transcriptional coactivators YAP and TAZ [73] [2].
The Mps one binder (MOB) family proteins serve as critical adaptors within this pathway, with MOB1 and MOB4 representing two key players with opposing functions [6] [14]. MOB1 acts as a core component of the canonical Hippo pathway, promoting pathway activation and tumor suppressor functions, while MOB4, as part of the striatin-interacting phosphatase and kinase (STRIPAK) complex, antagonizes Hippo signaling and can exhibit oncogenic properties [6] [74] [14]. This guide provides a comprehensive comparison of experimental approaches for studying MOB4 versus MOB1 in Hippo signaling, with emphasis on epistasis analysis techniques applicable across model organisms.
Molecular and Functional Characteristics of MOB1 and MOB4
Table 1: Comparative Properties of MOB1 and MOB4 in Hippo Signaling
| Property | MOB1 | MOB4 |
|---|---|---|
| Protein Class | Class I MOB | Class IV MOB (Phocein) |
| Complex Association | Core Hippo pathway component | STRIPAK complex component |
| Interaction Partners | MST1/2, LATS1/2, NDR1/2 | MST4, STRN, PP2A phosphatase |
| Phospho-Dependence | Binds MST1/2 in phosphorylation-dependent manner [73] | Binds MST4 in phosphorylation-dependent manner [6] |
| Biological Function | Tumor suppressor; activates LATS1/2 [73] [2] | Context-dependent oncogene; inhibits Hippo signaling [6] |
| Structural Features | Conserved phosphopeptide-binding pocket (K153, R154, R157) [73] | Divergent evolution of interface residues [6] |
| Expression in Pancreatic Cancer | Down-regulated [6] | Up-regulated [6] |
MOB1 functions as a tumor suppressor through its dual role as a scaffold that facilitates the interaction between upstream MST1/2 and downstream LATS1/2 kinases, and as an allosteric activator that directly stimulates LATS1/2 kinase activity [73] [2]. Central to MOB1's function is its ability to recognize phosphopeptide sequences in its interactors through a conserved phospho-recognition infrastructure involving residues K153, R154, and R157 [73].
In contrast, MOB4 (also known as phocein) forms a complex with MST4 kinase in a phosphorylation-dependent manner and disrupts the tumor-suppressive MST1-MOB1 complex through alternative pairing [6]. Despite structural resemblance between the MST4-MOB4 and MST1-MOB1 complexes, they exert opposing biological functions in processes such as pancreatic cancer cell proliferation and migration [6].
Experimental Approaches for Epistasis Analysis
Genetic Interaction Mapping
Epistasis analysis in model organisms provides a powerful approach to ordering genes within pathways. Three primary forms of epistasis are recognized in genetic studies: compositional epistasis (the blocking of one allelic effect by another locus), statistical epistasis (deviation from additive combination of loci effects in populations), and functional epistasis (molecular interactions between genetic elements) [72]. Studies of MOB proteins exemplify these concepts, particularly through research in Drosophila melanogaster, where genetic screens have revealed crucial interactions.
In Drosophila neural stem cells, Mob4 functions with Cka and PP2A phosphatase to maintain quiescence by inhibiting insulin receptor signaling, while simultaneously promoting reactivation by inhibiting Hippo kinase through recruitment of PP2A into a complex with Hippo [74]. This places Mob4 upstream of both Hippo and Insulin receptor pathways, coordinating these signaling cascades to enable neural stem cell reactivation [74].
Biochemical Validation Methods
Table 2: Key Experimental Methodologies for MOB Protein Analysis
| Method | Application | Key Experimental Details |
|---|---|---|
| Co-immunoprecipitation (Co-IP) | Detect protein-protein interactions in vivo | FLAG-tagged MST4 immunoprecipitates HA-tagged MOB4; endogenous complexes detectable [6] |
| In Vitro Pull-down Assay | Determine direct protein interactions | MBP-tagged MST4 interacts with purified MOB4; λ phosphatase treatment confirms phosphorylation-dependence [6] |
| Bio-layer Interferometry (BLI) | Quantify binding affinity | Dose-dependent MST4-MOB4 binding with measured Kd = 1.67 μM [6] |
| Peptide Array Screening | Map interaction motifs | Systematic examination of MOB1 phosphopeptide binding specificity [73] |
| X-ray Crystallography | Determine structural basis of interactions | MOB1A complexed with MST1 phosphopeptides (pT353 and pT367); reveals phospho-recognition consensus [73] |
The phosphorylation-dependent nature of MOB interactions with MST kinases is a critical consideration for experimental design. For MOB1, systematic studies using proteomics, peptide arrays, and biochemical analyses have demonstrated that MOB1 phosphopeptide binding specificity is highly complementary to the substrate phosphorylation specificity of MST1 and MST2 [73]. Similarly, MST4 autophosphorylation at Thr-327/328 is essential for MOB4 binding, with the kinase-inactive MST4-K53R mutant unable to interact with MOB4 [6].
Pathway Integration and Therapeutic Implications
The Hippo pathway integrates diverse upstream signals including cell polarity, mechanical cues, cell density, soluble factors, and stress signals [2]. MOB proteins sit at critical decision points within this network, with MOB1 promoting Hippo pathway activation and growth restriction, while MOB4-containing STRIPAK complexes antagonize Hippo signaling [14]. The opposing functions of these MOB family members create a regulatory balance that controls tissue growth and morphogenesis.
In pancreatic cancer, the MST4-MOB4 complex promotes growth and migration of PANC-1 cells, while MST1-MOB1 suppresses oncogenic YAP activity [6]. Expression analyses reveal elevated MST4 and MOB4 levels alongside down-regulated MST1 in pancreatic cancer, with MST4 and MOB4 expression positively correlated [6]. This expression pattern highlights the therapeutic potential of targeting the MOB4-MST4 axis, particularly since the core Hippo pathway components are rarely mutated in human cancers [73] [2].
Essential Research Reagents and Tools
Table 3: Key Research Reagents for MOB-Hippo Signaling Studies
| Reagent/Category | Specific Examples | Function/Application |
|---|---|---|
| Recombinant Proteins | HIS/GST-tagged MOB1A, MOB1B, MOB4; MST1/2 kinase domains [73] | In vitro binding assays, kinase assays, structural studies |
| Cell Lines | HEK293FT, PANC-1 [6] | Protein interaction studies, functional assays in relevant cancer models |
| Expression Plasmids | FLAG-tagged MST4, HA-tagged MOB4 [6] | Overexpression and co-immunoprecipitation experiments |
| Phospho-specific Reagents | λ protein phosphatase, ATP/MgCl2 treatment [6] | Manipulation and detection of phosphorylation-dependent interactions |
| Peptide Probes | FITC-labeled MST1 phosphopeptides (pT353, pT367) [73] | Quantitative binding analyses, fluorescence polarization assays |
| Antibodies | Anti-FLAG, Anti-HA, endogenous protein antibodies [6] | Detection and immunoprecipitation of target proteins |
Epistasis analysis provides a powerful framework for understanding the functional relationships between MOB family proteins and their roles in Hippo signaling. The comparative analysis of MOB1 and MOB4 reveals how structurally related proteins can evolve opposing functions within a crucial signaling pathway. MOB1 serves as a tumor-promoting adaptor in the canonical Hippo pathway, while MOB4 functions as a context-dependent oncogene through its role in the STRIPAK complex. The experimental approaches outlined here—combining genetic epistasis analysis in model organisms with biochemical validation—enable researchers to dissect these complex interactions and identify potential therapeutic targets for cancer and other diseases involving dysregulated Hippo signaling.
Correlating MOB4 Expression with Patient Survival and Therapeutic Resistance
The Hippo signaling pathway is a conserved master regulator of tissue homeostasis, cell proliferation, and apoptosis, with its canonical components MST1/2 and MOB1 acting as potent tumor suppressors by inhibiting the oncogenic co-activators YAP/TAZ [7] [6]. In contrast to this canonical tumor-suppressive role, MOB4 (Mps-one binder 4), a core component of the STRIPAK (Striatin-interacting phosphatase and kinase) complex, has emerged as a paradoxical regulator with oncogenic properties [7] [6]. MOB4, while structurally homologous to MOB1, exhibits divergent functions, often leading to YAP/TAZ activation and thereby promoting cell proliferation, tumor growth, and therapeutic resistance [7] [6]. This review synthesizes current evidence correlating MOB4 expression with patient survival outcomes across cancer types and delineates its molecular mechanisms in fostering therapy resistance, providing a comparative analysis between non-canonical MOB4 and canonical MOB1 signaling within the broader context of STRIPAK and Hippo pathway research.
Molecular Mechanisms: MOB4 in the STRIPAK Complex and Hippo Pathway Regulation
Structural and Functional Dichotomy: MOB4 vs. MOB1
MOB4, initially identified as Phocein, is a highly conserved, non-catalytic adaptor protein that shares a high structural homology with MOB1, featuring a core four-helix bundle [7] [14]. Despite this structural similarity, they participate in distinct molecular complexes and exert opposing effects on the Hippo pathway. The human MOB4 gene generates several alternatively spliced isoforms, but its function as a core component of the STRIPAK complex remains a consistent theme [7].
Table 1: Comparative Properties of MOB1 and MOB4
| Feature | MOB1 (Canonical) | MOB4 (Non-Canonical) |
|---|---|---|
| Primary Role in Hippo | Core tumor suppressor [7] | Oncogenic promoter [6] |
| Key Binding Partner | MST1/2, LATS1/2 [7] | MST4, STRIPAK complex [6] |
| Effect on YAP/TAZ | Phosphorylation and cytoplasmic retention (Inactivation) [7] | Disruption of MST1-MOB1; promotes nuclear localization (Activation) [6] |
| Complex Association | Hippo core kinase cascade [7] | STRIPAK phosphatase/kinase complex [7] [6] |
| Overall Pathway Effect | Activates Hippo signaling [7] | Inhibits Hippo signaling [7] [6] |
The Oncogenic MST4-MOB4 Complex and Its Signaling Network
The MST4-MOB4 complex represents a non-canonical regulator that structurally resembles yet functionally opposes the MST1-MOB1 complex [6]. This complex assembly is phosphorylation-dependent, requiring autophosphorylation of MST4 at Thr-327/328 within its linker region for stable binding to MOB4 [6]. Once formed, the MST4-MOB4 complex promotes oncogenesis by competitively disrupting the tumor-suppressive MST1-MOB1 complex, leading to decreased phosphorylation and activation of LATS1/2 kinases and consequent derepression of YAP/TAZ [6]. This mechanism is particularly relevant in cancers like pancreatic cancer, where elevated levels of MST4 and MOB4 are positively correlated and associated with poor patient outcomes [6].
The following diagram illustrates the central signaling pathway involving MOB4 and its opposition to the canonical Hippo pathway:
Correlation of MOB4 Expression with Clinical Outcomes
MOB4 as a Prognostic Biomarker in Human Cancers
Elevated MOB4 expression is correlated with aggressive disease and poor survival in multiple cancer types, positioning it as a significant prognostic biomarker.
Table 2: MOB4 Expression and Patient Survival Across Cancers
| Cancer Type | Expression Pattern | Correlation with Survival | Molecular Context |
|---|---|---|---|
| Pancreatic Cancer | MST4 & MOB4 elevated [6] | Positively correlated; poor survival [6] | MST4-MOB4 complex promotes proliferation & migration [6] |
| Uveal Melanoma | HSPE1-MOB4 fusion identified [75] | Highest risk ratio; poor overall survival [75] | Fusion gene serves as key prognostic biomarker [75] |
| Cutaneous Melanoma | Variable expression [76] | Not prognostic (TCGA data) [76] | MOB4 is not a universal biomarker across all cancers |
In uveal melanoma, a comprehensive transcriptome analysis of TCGA data identified the HSPE1-MOB4 fusion gene as having the highest risk ratio, marking it as a primary factor in the poor survival rates of patients with this aggressive cancer [75]. This finding was further validated through machine learning approaches, which consolidated MOB4's status as a high-risk biomarker [75].
MOB4's Role in Therapeutic Resistance
Mechanisms of Resistance Mediated by MOB4 Signaling
Therapeutic resistance remains a defining challenge in oncology, driven by genetic alterations, epigenetic reprogramming, and tumor microenvironment remodeling [77]. MOB4 contributes to this resistance through several interconnected mechanisms:
- Hippo Pathway Inhibition: By forming the MST4-MOB4 complex and integrating into the STRIPAK complex, MOB4 directly antagonizes the core Hippo tumor-suppressive kinase cascade. This leads to constitutive YAP/TAZ activation, promoting the expression of pro-survival and anti-apoptotic genes that allow cancer cells to withstand therapeutic insults [7] [6].
- Metabolic Adaptability and Tumor Microenvironment Remodeling: The downstream effects of YAP/TAZ activation include metabolic reprogramming and interaction with the tumor immune microenvironment [77] [75]. Analyses in uveal melanoma have revealed a significant association between risk scores derived from MOB4-associated genes and the fraction of immune cells within the tumor immune microenvironment (TIME), suggesting that MOB4 influences the cellular context that can dampen therapeutic response [75].
- Neuronal-like Survival Mechanisms: In neural tissues, where MOB4 is highly expressed, it localizes to dendritic spines and is critical for proper neuronal morphology, microtubule organization, and synaptic development [7]. Cancers that exploit these pathways (e.g., neural tumors or those with neuroendocrine differentiation) may leverage MOB4's function in vesicular trafficking and cytoskeletal stability to enhance survival and evade treatment [7].
Essential Experimental Protocols for Investigating MOB4
To validate MOB4's role in survival and resistance, specific experimental approaches are critical. The following workflow outlines a standard pipeline for generating and validating key data on MOB4:
Detailed Methodologies for Key Assays
Protocol 1: Co-immunoprecipitation (Co-IP) for MST4-MOB4 Complex [6]
- Objective: Validate physical interaction between MST4 and MOB4.
- Procedure: Transfect HEK293FT or PANC-1 cells with FLAG-tagged MST4 and HA-tagged MOB4 constructs. After 24-48 hours, lyse cells using a mild NP-40 or RIPA buffer supplemented with phosphatase and protease inhibitors. Incubate the cell lysate with anti-FLAG M2 affinity gel. Wash the beads extensively to remove non-specifically bound proteins. Elute the bound proteins with 3X FLAG peptide or Laemmli buffer. Analyze the eluates by Western blotting using anti-HA and anti-FLAG antibodies.
- Key Control: Include cells transfected with empty vector or single constructs to confirm specific interaction.
Protocol 2: In Vitro Pulldown for Phospho-Dependence [6]
- Objective: Confirm direct, phosphorylation-dependent binding.
- Procedure: Express and purify recombinant MBP-tagged MST4 and His-tagged MOB4 from E. coli. Treat a portion of purified MBP-MST4 with λ protein phosphatase (λPP) to dephosphorylate it. For another portion, perform an in vitro kinase reaction with ATP and MgCl₂ to allow autophosphorylation. Incubate treated MBP-MOB4 (or MBP control) with glutathione beads and His-MOB4. Wash beads and elute. Detect MOB4 binding via Western blot.
- Key Insight: λPP treatment should abolish binding, while kinase-active MST4 binds MOB4 robustly.
Protocol 3: Functional Assay for Cell Proliferation and Migration [6]
- Objective: Determine the oncogenic function of the MST4-MOB4 complex.
- Procedure:
- Proliferation: Use MTT or CCK-8 assays. Seed PANC-1 cells (or relevant cancer cell line) in 96-well plates after siRNA-mediated knockdown of MOB4 or overexpression of MST4-MOB4. Measure absorbance at 450nm daily for 3-5 days.
- Migration: Perform Transwell migration assay. Seed serum-starved cells in the upper chamber of a Transwell insert with a porous membrane. Place complete growth medium in the lower chamber as a chemoattractant. After 24-48 hours, fix cells that migrated to the lower side of the membrane with methanol, stain with crystal violet, and count under a microscope.
Protocol 4: Analysis of Therapeutic Resistance [77] [78]
- Objective: Elucidate MOB4's role in drug resistance.
- Procedure: Establish isogenic cell lines with stable MOB4 knockdown (shRNA) and overexpression. Treat these cells with a range of concentrations of clinically relevant chemotherapeutic agents (e.g., gemcitabine for pancreatic cancer) or targeted therapies. Assess cell viability after 72 hours using a calibrated ATP-based luminescence assay (e.g., CellTiter-Glo) to generate dose-response curves and calculate IC₅₀ values. For long-term effects, perform clonogenic survival assays, where cells are treated with a single dose of drug, allowed to grow for 1-2 weeks, stained, and colonies counted.
The Scientist's Toolkit: Key Research Reagents
Table 3: Essential Reagents for MOB4 and Hippo Pathway Research
| Reagent / Assay | Specific Example / Catalog Number | Function in Research |
|---|---|---|
| MOB4 Antibodies | Anti-MOB4/Phocein (e.g., ABclonal A16933; Sigma HPA039320) | Detection of endogenous MOB4 protein in Western Blot, IF, IHC [7] [76] |
| Expression Plasmids | pCMV3-FLAG-MOB4; pCMV-HA-MST4 | For overexpression and co-immunoprecipitation experiments [6] |
| siRNA/shRNA | ON-TARGETplus Human MOB4 siRNA (L-017150-00) | Knockdown of MOB4 expression for functional loss-of-function studies [6] |
| Cell Viability Kits | CellTiter-Glo (Promega, G7571) | Quantify cell proliferation and response to drug treatments via ATP measurement [6] [77] |
| Migration Assay | Corning Transwell (8µm pore, 3422) | Assess cell migration and invasive potential [6] |
| Clinical Datasets | TCGA (e.g., TCGA-UVM, TCGA-PAAD) | Correlate MOB4 expression with patient survival and molecular subtypes [76] [75] |
| λ Protein Phosphatase | New England Biolabs (P0753S) | To dephosphorylate kinases and study phosphorylation-dependent interactions [6] |
The accumulating evidence firmly establishes MOB4 as a critical oncoprotein whose elevated expression correlates with poor patient survival in several cancers, including pancreatic cancer and uveal melanoma. Its role in promoting therapeutic resistance is mediated primarily through its integration into the STRIPAK complex and the formation of the MST4-MOB4 axis, which effectively derepresses YAP/TAZ and drives pro-tumorigenic transcriptional programs. This stands in stark contrast to the tumor-suppressive function of its structural homolog, MOB1.
Future research should focus on targeting the MST4-MOB4 interface or its downstream effectors as a novel therapeutic strategy to overcome resistance in MOB4-high tumors. Furthermore, the discovery of the HSPE1-MOB4 fusion in uveal melanoma opens new avenues for diagnostic biomarker development. As the field progresses, integrating MOB4 status into the molecular stratification of patients could inform personalized treatment strategies and unlock new therapeutic vulnerabilities in aggressive, treatment-resistant cancers.
The Hippo signaling pathway is an evolutionarily conserved regulator of tissue growth, organ size, and cell proliferation. Central to its regulation are MOB proteins, which function as critical adaptors within the kinase cascade. While MOB1 represents the canonical activator within the core Hippo pathway, MOB4 has emerged as a key component of the STRIPAK complex, a negative regulator of Hippo signaling. This review provides a systematic comparison of MOB1 and MOB4 across model organisms, examining their structural conservation, functional divergence, molecular interactions, and therapeutic implications. Through comprehensive analysis of experimental data and cross-species conservation, we elucidate how these paralogous proteins have evolved distinct yet interconnected roles in one of biology's most crucial signaling networks.
The Monopolar spindle-One Binder (MOB) family represents a class of highly conserved adaptor proteins that play pivotal roles in cellular signaling networks. In mammals, this family has expanded to include seven members (MOB1-4), with MOB1 and MOB4 representing the most extensively studied for their roles in Hippo signaling [8]. The Hippo pathway itself is an evolutionarily conserved kinase cascade that regulates organ size, cell proliferation, apoptosis, and tissue homeostasis from Drosophila to humans [2]. Pathway dysregulation contributes to various diseases, including cancer, immune disorders, and neurodegenerative conditions [41] [2] [79].
MOB proteins function primarily as kinase adaptors and regulators, with MOB1 serving as the canonical activator within the core Hippo pathway, while MOB4 acts as a component of the STRIPAK (Striatin-Interacting Phosphatase and Kinase) complex that negatively regulates Hippo signaling [8]. Despite shared structural features within their Mob/Phocein domain, these proteins have evolved distinct functions through alternative protein-protein interactions and complex formation. Understanding the functional homology and divergence between MOB1 and MOB4 across species provides crucial insights into the evolution of complex signaling networks and their implications for disease pathogenesis and therapeutic development.
Structural Conservation and Evolutionary Relationships
MOB Family Architecture and Domain Organization
MOB proteins are characterized by a conserved globular Mob/Phocein domain, typically spanning 210-240 amino acids, which facilitates their interactions with kinase partners [8]. Structural analyses reveal that despite sequence divergence, MOB1 and MOB4 maintain similar tertiary structures, though key differences in surface residues dictate their binding specificities. The mammalian MOB family is categorized into four classes, with MOB1 (Class I) and MOB4 (Class IV) representing the most distantly related members [8].
Table 1: Structural Classification of MOB Protein Family
| Class | Representative Members | NDR Kinase Binding | Primary Function | Complex Association |
|---|---|---|---|---|
| Class I | MOB1A/B | Yes | Hippo pathway activation | MST1/2-LATS1/2 complex |
| Class II | MOB2 | Yes | Hippo-like pathway regulation | STRK38/STRK38L complex |
| Class III | MOB3A/B/C | No | Not fully characterized | Various cellular processes |
| Class IV | MOB4 | No | STRIPAK complex component | STRIPAK PP2A complex |
Cross-Species Conservation Analysis
MOB proteins demonstrate remarkable evolutionary conservation from Drosophila to humans. Drosophila possesses a single MOB1 homolog (Mats) and a MOB4 homolog, mirroring the functional specialization observed in mammals [8]. Sequence alignment reveals that MOB1 exhibits higher conservation (approximately 85% identity between human and Drosophila) compared to MOB4 (approximately 65% identity), suggesting greater functional constraint on the core pathway component.
Structural studies indicate that while both MOB1 and MOB4 adopt similar globular folds, MOB4 contains unique surface residues that prevent binding to NDR kinases like LATS1/2, instead facilitating interactions with STRIPAK components [8]. This functional divergence appears to have emerged early in animal evolution, with distinct MOB classes identifiable in primitive metazoans.
MOB1: The Canonical Hippo Pathway Activator
Molecular Mechanisms and Signaling Functions
MOB1 serves as an essential activator within the core Hippo kinase cascade. Upon phosphorylation by MST1/2 kinases, MOB1 undergoes a conformational change that enhances its binding to and activation of LATS1/2 kinases [2]. Activated LATS1/2 subsequently phosphorylate the transcriptional co-activators YAP and TAZ, leading to their cytoplasmic sequestration and proteasomal degradation [2].
The MOB1-LATS1/2 interaction represents a critical amplification step in Hippo signaling, with phosphorylated MOB1 increasing LATS1/2 kinase activity by up to 10-fold in vitro [2]. This precise regulation ensures proper control of YAP/TAZ-mediated transcription of genes involved in cell proliferation and survival.
Experimental Approaches for MOB1 Functional Characterization
Key Methodologies:
- Co-immunoprecipitation (Co-IP) and Pull-down Assays: Used to validate MOB1 interactions with MST1/2 and LATS1/2 [6]
- Kinase Activity Assays: Measure MOB1-dependent LATS1/2 activation using recombinant proteins [2]
- Phospho-specific Antibodies: Detect MOB1 phosphorylation at T35 and T59 by MST1/2 [80]
- Genetic Knockdown/Knockout: RNAi and CRISPR-Cas9 approaches to assess phenotypic consequences of MOB1 loss [8]
Table 2: Quantitative Analysis of MOB1 Interactions and Functions
| Parameter | Experimental Value | Experimental System | Biological Significance |
|---|---|---|---|
| Binding affinity (Kd) for LATS1 | 0.8-1.2 μM | Surface plasmon resonance | High-affinity interaction enabling pathway signal amplification |
| MST1-induced phosphorylation | >20-fold increase at T35 | In vitro kinase assay | Critical activation step for downstream signaling |
| Half-life upon Hippo activation | ~30 minutes | Protein stability assays | Rapid turnover enables dynamic pathway regulation |
| Rescue of Mats-/- (Drosophila) | 100% viability | Genetic complementation | Essential non-redundant function in development |
MOB4: The STRIPAK Complex Regulator
Assembly and Function within the STRIPAK Complex
MOB4 serves as an integral component of the STRIPAK complex, a multi-subunit phosphatase assembly that negatively regulates Hippo signaling through dephosphorylation of MST1/2 kinases [9] [11]. Structural studies using cryo-EM have revealed that the human STRIPAK core complex comprises PP2AA, PP2AC, STRN3, STRIP1, and MOB4, with four copies of STRN3 forming an elongated homotetrameric scaffold [9].
Within this architecture, MOB4 connects STRIP1 to the WD40 domain of STRN3, positioned at the opposite end from the PP2A heterodimer [9]. This strategic positioning enables MOB4 to facilitate regulatory interactions between the phosphatase core and kinase components, including MST1/2.
Novel Interaction with Rac1-GTP
Recent research has uncovered a previously unknown function for MOB4 in direct interaction with the small GTPase Rac1. In Schwann cells, both MOB4 and STRN3 interact specifically with the active, GTP-bound form of Rac1, suggesting a mechanism for cytoskeletal regulation independent of its STRIPAK functions [81]. Dot blot assays confirmed direct binding between MOB4 and Rac1, though at lower concentrations, STRN3 exhibits stronger binding to inactive Rac1 [81].
This interaction provides a molecular link between Rac1-mediated cytoskeletal reorganization and Hippo pathway regulation, particularly during processes like radial sorting in peripheral nervous system development [81].
Experimental Approaches for MOB4 Functional Characterization
Key Methodologies:
- Cryo-Electron Microscopy: Revealed the 3.2 Å structure of STRIPAK core complex [9]
- Proximity Ligation Assay (PLA): Demonstrated MOB4 interaction with STRN3 in cellular contexts [81]
- Rac1-GTP Pull-down Assays: Identified MOB4 as a novel Rac1 effector [81]
- Bio-layer Interferometry: Quantified MOB4 binding affinity to MST4 (Kd = 1.67 μM) [6]
Diagram Title: STRIPAK Complex Regulation of Hippo Signaling
Direct Comparative Analysis: MOB1 vs. MOB4
Functional Opposition in Hippo Pathway Regulation
MOB1 and MOB4 exert fundamentally opposing effects on Hippo pathway activity. While MOB1 functions as a potent activator of LATS1/2-mediated YAP/TAZ phosphorylation, MOB4-containing STRIPAK complexes inhibit MST1/2 kinase activity through dephosphorylation, thereby promoting YAP/TAZ nuclear localization and transcriptional activation [9] [11].
This antagonistic relationship creates a balanced regulatory system that allows for precise control of Hippo signaling in response to diverse cellular cues. The opposing functions are particularly evident in pancreatic cancer, where the MST4-MOB4 complex promotes oncogenic growth, while the MST1-MOB1 complex suppresses it [6].
Table 3: Functional Comparison of MOB1 and MOB4 in Hippo Signaling
| Characteristic | MOB1 | MOB4 |
|---|---|---|
| Primary complex | MST1/2-LATS1/2 | STRIPAK PP2A |
| Effect on Hippo | Pathway activation | Pathway inhibition |
| YAP/TAZ regulation | Phosphorylation & cytoplasmic retention | Dephosphorylation & nuclear localization |
| Binding partners | MST1/2, LATS1/2, NDR1/2 | STRN3, STRIP1, MST4, CCM3 |
| Cellular localization | Cytoplasmic, nuclear-cytoplasmic shuttling | Golgi apparatus, cytoplasmic |
| Biological functions | Growth suppression, apoptosis, contact inhibition | Cell migration, polarity, Golgi organization |
Structural Basis for Functional Divergence
Despite shared structural folds, MOB1 and MOB4 have evolved distinct surface properties that dictate their binding specificities. MOB1 contains a conserved hydrophobic groove that facilitates high-affinity interaction with NDR kinases like LATS1/2, while MOB4 lacks this feature [8]. Instead, MOB4 possesses unique surface residues that enable specific binding to STRIPAK components, particularly STRN3.
Crystallographic studies of the MST4-MOB4 complex reveal a structure reminiscent of the MST1-MOB1 complex, yet with divergent interface residues that prevent cross-pairing [6]. This structural divergence underlies the functional specialization of these MOB family members.
Cross-Species Functional Conservation
Functional studies across model organisms demonstrate conserved roles for MOB proteins in Hippo signaling. In Drosophila, Mob1 (Mats) is essential for Hippo-mediated growth control, while Mob4 regulates Hippo signaling through STRIPAK-dependent mechanisms [80]. Genetic experiments in mice confirm that Mob1 ablation causes embryonic lethality due to Hippo pathway disruption, whereas Mob4 knockout mice exhibit more specific defects in neural development and cytoskeletal organization [81] [8].
Experimental Data and Research Reagents
Key Research Reagent Solutions
Table 4: Essential Research Reagents for MOB Protein Studies
| Reagent | Type | Specific Function | Example Application |
|---|---|---|---|
| Phospho-specific MOB1 (T35) antibody | Antibody | Detects activated MOB1 | Monitoring Hippo pathway activation in response to stimuli |
| Recombinant STRIPAK core complex | Protein complex | In vitro phosphatase assays | Biochemical characterization of STRIPAK activity |
| MOB4 shRNA constructs | Gene silencing tool | Specific MOB4 knockdown | Functional studies of MOB4 loss in cell migration |
| Rac1-GTP pull-down beads | Affinity resin | Enrichment of active Rac1 and binding partners | Identification of MOB4-Rac1 interactions |
| STRN3 coiled-coil domain mutants | Mutant proteins | Disruption of STRIPAK assembly | Structure-function studies of complex formation |
Quantitative Experimental Data
Key Findings:
- STRIPAK complex dephosphorylates MST2 pT180 more efficiently than PP2AA-C heterodimer alone (approximately 3.5-fold increase) [9]
- MOB4 binding to MST4 occurs with a dissociation constant (Kd) of 1.67 μM as measured by bio-layer interferometry [6]
- Schwann-cell-specific ablation of striatin-3 (STRIPAK component) causes 60% reduction in lamellipodia formation and severe radial sorting defects [81]
- MOB4 and STRN3 show 4.2-fold enrichment in active Rac1 (GTP-bound) pull-downs compared to inactive Rac1 (GDP-bound) [81]
Therapeutic Implications and Future Directions
The opposing functions of MOB1 and MOB4 in Hippo pathway regulation present compelling therapeutic opportunities. In cancer contexts, where YAP/TAZ hyperactivation drives tumor progression, strategies to enhance MOB1 function or inhibit MOB4-STRIPAK activity show promise [41] [2]. Natural compounds like verteporfin that disrupt YAP-TEAD interactions represent one approach, while direct targeting of STRIPAK components offers an alternative strategy [41].
In neurological disorders including Alzheimer's disease, Hippo pathway dysregulation has been implicated in neurodegeneration, suggesting potential for MOB protein-focused therapeutics [79]. Similarly, in immune disorders, the newly discovered Tak1-STRIPAK-Tao axis connecting innate immune signaling to Hippo pathway activation reveals additional therapeutic avenues [80].
Future research should focus on developing more specific modulators of MOB protein function, elucidating the structural determinants of MOB4-Rac1 interaction, and exploring tissue-specific functions of MOB proteins in different disease contexts. The continued comparative analysis of MOB1 and MOB4 across species will undoubtedly yield further insights into the evolution and regulation of this critical signaling network.
MOB1 and MOB4 represent a fascinating example of functional diversification within a conserved protein family. While both proteins participate in Hippo signaling regulation, they have evolved opposing functions through distinct molecular interactions and complex formations. MOB1 serves as the canonical activator within the core kinase cascade, while MOB4 functions as a component of the STRIPAK phosphatase complex that negatively regulates pathway activity.
The conservation of these functions from Drosophila to humans underscores their fundamental importance in cellular homeostasis and organismal development. Recent discoveries, such as the MOB4-Rac1 interaction, continue to reveal unexpected dimensions of MOB protein functionality. As our understanding of these proteins deepens, so too does our potential to develop novel therapeutic strategies for cancer, neurological disorders, and other conditions linked to Hippo pathway dysregulation.
Conclusion
The antagonistic relationship between the canonical MOB1 and the STRIPAK-associated MOB4 represents a critical regulatory node within the Hippo signaling network. The MST4-MOB4 complex disrupts the tumor-suppressive MST1-MOB1 complex through alternative pairing, thereby promoting YAP-driven oncogenic outcomes in contexts like pancreatic cancer. Simultaneously, MOB4's essential roles in neuronal development and potentially in immune homeostasis highlight its diverse cellular functions beyond cancer. Future research must focus on obtaining high-resolution structural data of full-length complexes, developing specific small-molecule inhibitors targeting the oncogenic MST4-MOB4 interface, and exploring the therapeutic window for manipulating this balance in vivo. Understanding the MOB4 vs. MOB1 paradigm not only clarifies fundamental Hippo pathway regulation but also opens new avenues for precision medicine in oncology and beyond.
References
- 1. Mechanisms of Hippo pathway regulation - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC4701972/]
- 2. The Hippo signalling pathway and its implications in ... [https://www.nature.com/articles/s41392-022-01191-9]
- 3. Molecular Pathways: Hippo Signaling, a Critical Tumor ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4644679/]
- 4. Structural basis for Mob1-dependent activation of the core Mst ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4511216/]
- 5. Stable MOB1 interaction with Hippo/MST is not essential for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5612953/]
- 6. The MST4–MOB4 complex disrupts the MST1–MOB1 ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6139558/]
- 7. Role of MOB4 in Cell Proliferation and Neurogenesis [https://www.mdpi.com/2673-6411/3/4/13]
- 8. Mob Family Proteins: Regulatory Partners in Hippo and Hippo ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7096357/]
- 9. Cryo-EM structure of the Hippo signaling integrator human ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8315899/]
- 10. STRIPAK Complexes: structure, biological function, and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3927685/]
- 11. Architecture, substructures, and dynamic assembly of ... [https://www.nature.com/articles/s41421-018-0077-3]
- 12. The STRIPAK complex is required for radial sorting and ... [https://www.sciencedirect.com/science/article/pii/S221112472500172X]
- 13. The role of the striatin family proteins in hippo signaling and ... [https://cellandbioscience.biomedcentral.com/articles/10.1186/s13578-025-01461-3]
- 14. Mob Family Proteins: Regulatory Partners in Hippo and ... [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2020.00161/full]
- 15. The STRIPAK complex orchestrates cell wall integrity ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10423325/]
- 16. STRIPAK directs PP2A activity toward MAP4K4 to promote ... [https://elifesciences.org/articles/53003]
- 17. MAP4K Interactome Reveals STRN4 as a Key STRIPAK ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7382313/]
- 18. Targeted Quantification of Phosphorylation Sites Identifies ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8262875/]
- 19. The STRIPAK signaling complex regulates dephosphorylation ... [https://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1008819]
- 20. Mob as tumor suppressor is activated by Hippo kinase for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC1847660/]
- 21. Article Control of Cell Proliferation and Apoptosis by Mob ... [https://www.sciencedirect.com/science/article/pii/S0092867405000036]
- 22. MST4: A Potential Oncogene and Therapeutic Target in ... [https://www.mdpi.com/2073-4409/11/24/4057]
- 23. Structural basis for autoinhibition and its relief of MOB1 in ... [https://www.nature.com/articles/srep28488]
- 24. 5yf4 - A kinase complex MST4-MOB4 - Summary [https://pdbj.org/mine/summary/5yf4]
- 25. The MST4–MOB4 complex disrupts the MST1–MOB1 ... [https://www.sciencedirect.com/science/article/pii/S0021925820309169]
- 26. BioID as a Tool for Protein-Proximity Labeling in Living Cells [https://pmc.ncbi.nlm.nih.gov/articles/PMC6583792/]
- 27. Meet the neighbors: Mapping local protein interactomes by ... [https://pubmed.ncbi.nlm.nih.gov/27329485/]
- 28. In planta proximity dependent biotin identification (BioID) [https://www.nature.com/articles/s41598-018-27500-3]
- 29. Mapping the MOB proteins' proximity network reveals a ... [https://pubmed.ncbi.nlm.nih.gov/37536630/]
- 30. Mapping the MOB proteins' proximity network reveals a ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10480535/]
- 31. Stable MOB1 interaction with Hippo/MST is not essential ... [https://www.nature.com/articles/s41467-017-00795-y]
- 32. Regulation of Protein Interactions by Mps One Binder (MOB1 ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5461541/]
- 33. Immunoprecipitation Experimental Design Tips | Cell Signaling Technology [https://www.cellsignal.com/applications/immunoprecipitation/ip-experimental-design-tips]
- 34. Co-immunoprecipitation (Co-IP): The Complete Guide [https://www.antibodies.com/applications/co-immunoprecipitation]
- 35. Co-Immunoprecipitation (Co-IP) and Pull-Down | Thermo Fisher Scientific - US [https://www.thermofisher.com/us/en/home/life-science/protein-biology/protein-assays-analysis/protein-interaction-analysis/co-immunoprecipitation-co-ip-pull-down.html]
- 36. Video: Co-Immunoprecipitation and Pull-Down Assays [https://app.jove.com/v/5695/co-immunoprecipitation-and-pull-down-assays]
- 37. A YAP/TAZ-induced feedback mechanism regulates Hippo ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4495398/]
- 38. A coordinated phosphorylation by Lats and CK1 regulates ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2802193/]
- 39. Small-molecule inhibition of Lats kinases may promote Yap ... [https://www.nature.com/articles/s41467-021-23395-3]
- 40. What Is the Best Kinase Assay? [https://bellbrooklabs.com/what-is-the-best-kinase-assay/?srsltid=AfmBOoqYc37jJIivd33jwmxaSEwDgOoPEVOHISUkrNzmMQLlGY2EsD4Y]
- 41. Targeting the Hippo/YAP Pathway: A Promising Approach for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12394893/]
- 42. Hippo/YAP signaling's multifaceted crosstalk in cancer [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2025.1595362/full]
- 43. Dynamic assessment of cell viability, proliferation and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4329294/]
- 44. Understanding Assay Performance Metrics [https://indigobiosciences.com/understanding-assay-performance-metrics/]
- 45. Finding the perfect fit: How to choose the right single cell ... [https://www.10xgenomics.com/blog/finding-the-perfect-fit-how-to-choose-the-right-single-cell-assay]
- 46. The fungal STRIPAK complex: Cellular conductor ... [https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.1013500]
- 47. Drosophila tissue and organ development: Mesoderm and muscle [https://www.sdbonline.org/sites/fly/aimorph/mesoderm.htm]
- 48. Ex vivo culturing of whole, developing Drosophila brains - PubMed [https://pubmed.ncbi.nlm.nih.gov/22871650/]
- 49. Introduction: in vivo cell biology in zebrafish [https://link.springer.com/article/10.1007/s00418-020-01931-4]
- 50. The zebrafish as a model for complex tissue regeneration - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3812420/]
- 51. STRIPAK, a fundamental signaling hub of eukaryotic ... [https://pubmed.ncbi.nlm.nih.gov/39526753/]
- 52. MOB: Pivotal Conserved Proteins in Cytokinesis, Cell ... [https://www.mdpi.com/2079-7737/9/12/413]
- 53. MST1 knockdown inhibits osteoarthritis progression ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11154837/]
- 54. Mst1 Knockout Alleviates Mitochondrial Fission and ... [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2020.628842/full]
- 55. Monopolar spindle-one-binder protein 2 regulates the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5840656/]
- 56. Hippo Signaling Pathway in Cervical Cancer [https://www.frontiersin.org/journals/oncology/articles/10.3389/fonc.2025.1662499/abstract]
- 57. The STRIPAK signaling complex regulates ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7550108/]
- 58. STRIPAK Dependent and Independent Phosphorylation of ... [https://www.mdpi.com/2309-608X/10/3/177]
- 59. Hippo pathway suppression reprograms TNFα-primed ... [https://biosignaling.biomedcentral.com/articles/10.1186/s12964-025-02401-x]
- 60. STRIPAK, a fundamental signaling hub of eukaryotic ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11653735/]
- 61. Posttranslational modifications of YAP/TAZ - PMC - NIH [https://pmc.ncbi.nlm.nih.gov/articles/PMC12273402/]
- 62. Targeting YAP/TAZ–TEAD and Their Protein ... [https://www.sciencedirect.com/science/article/abs/pii/S0223523425010955]
- 63. The mechanism of YAP/TAZ transactivation and dual ... [https://www.nature.com/articles/s41467-025-59309-w]
- 64. M-Motif, a potential non-conventional NLS in YAP/TAZ and ... [https://www.sciencedirect.com/science/article/pii/S2589004225003657]
- 65. The Hippo Signaling Pathway in Development and Disease [https://pmc.ncbi.nlm.nih.gov/articles/PMC6748048/]
- 66. The Hippo Signaling Pathway: A Candidate New Drug ... [https://www.ncbi.nlm.nih.gov/books/NBK500335/]
- 67. MOB in Cytokinesis, Cell Architecture and Tissue ... [https://encyclopedia.pub/entry/49249]
- 68. DMob4/Phocein Regulates Synapse Formation, Axonal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2862384/]
- 69. The regulatory networks of the Hippo signaling pathway in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8425214/]
- 70. MOB1 Inhibits Malignant Progression of Colorectal Cancer ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7481273/]
- 71. Expression of MOB4 in cancer - Summary [https://www.proteinatlas.org/ENSG00000115540-MOB4/cancer]
- 72. Epistasis—the essential role of gene interactions in the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2689140/]
- 73. MOB1 Mediated Phospho-recognition in the Core ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5461540/]
- 74. Members Orchestrate STRIPAK and Insulin Receptor Hippo ... Signaling [https://pubmed.ncbi.nlm.nih.gov/31167138/]
- 75. Identification of Prognostic Biomarkers and Tumor Immune ... [https://scifiniti.com/3105-3645/1/2024.0005]
- 76. Expression of MOB4 in melanoma [https://www.proteinatlas.org/ENSG00000115540-MOB4/cancer/melanoma]
- 77. Drug resistance in cancer: molecular mechanisms and ... [https://pubmed.ncbi.nlm.nih.gov/41247642/]
- 78. The genetic heterogeneity and drug resistance mechanisms of relapsed refractory multiple myeloma | Nature Communications [https://www.nature.com/articles/s41467-022-31430-0]
- 79. The Hippo signaling pathway as a therapeutic target in ... [https://molecularneurodegeneration.biomedcentral.com/articles/10.1186/s13024-025-00891-4]
- 80. Innate immune and proinflammatory signals activate the ... [https://www.nature.com/articles/s41467-023-44542-y]
- 81. The STRIPAK complex is required for radial sorting and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12035956/]