Magnesium Concentration in PCR: A Comprehensive Guide to Optimizing Reaction Fidelity and Specificity
This article provides a systematic evaluation of how magnesium ion (Mg²⁺) concentration fundamentally influences the fidelity and efficiency of the Polymerase Chain Reaction (PCR).
Magnesium Concentration in PCR: A Comprehensive Guide to Optimizing Reaction Fidelity and Specificity
Abstract
This article provides a systematic evaluation of how magnesium ion (Mg²⁺) concentration fundamentally influences the fidelity and efficiency of the Polymerase Chain Reaction (PCR). Tailored for researchers, scientists, and drug development professionals, the content explores the biochemical role of Mg²⁺ as an essential polymerase cofactor, details practical strategies for its optimization across various template types and applications, and presents advanced troubleshooting methodologies. By synthesizing foundational knowledge with applied protocols and comparative validation techniques, this guide aims to equip practitioners with the evidence-based framework necessary to enhance the accuracy and reproducibility of their molecular assays, which is critical for applications in diagnostics, cloning, and sequencing.
The Fundamental Role of Magnesium Ions in PCR Fidelity and DNA Polymerase Activity
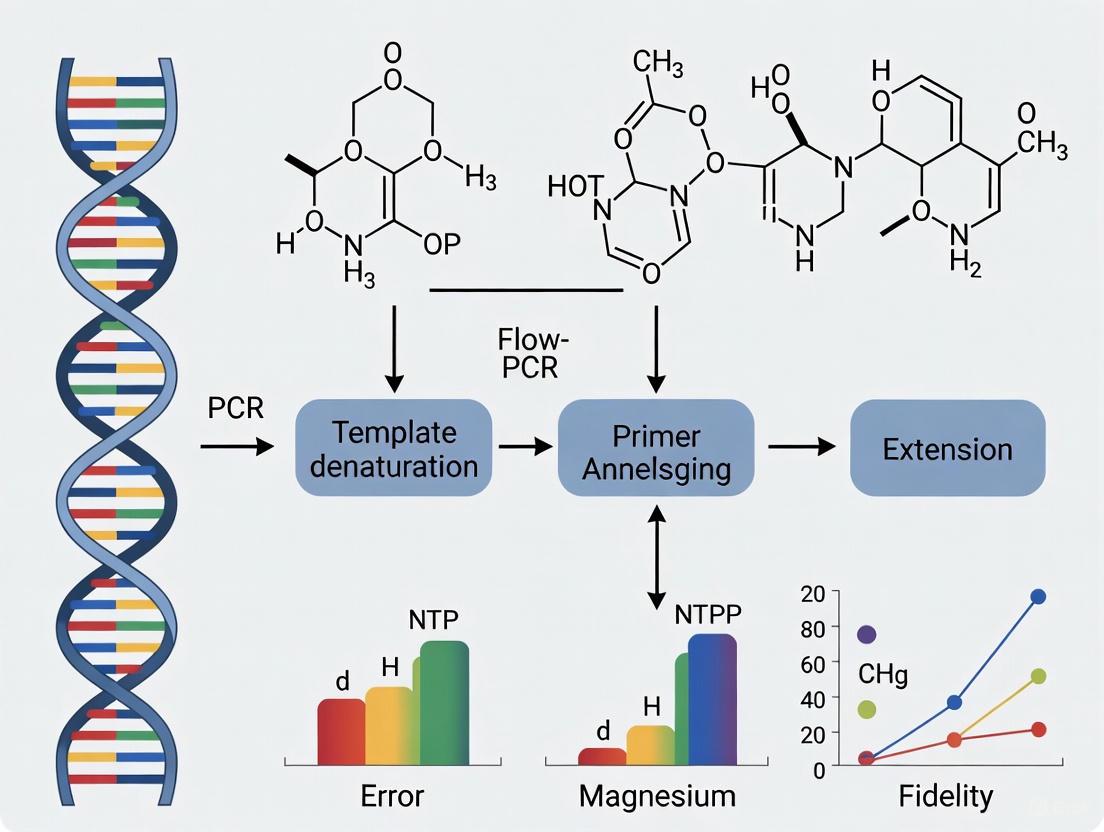
Magnesium ions (Mg²⁺) are indispensable for the catalytic activity of DNA polymerases in the Polymerase Chain Reaction (PCR), serving as a critical cofactor that directly facilitates the nucleophilic attack during phosphodiester bond formation. The concentration of Mg²⁺ in the reaction buffer is a pivotal determinant of PCR success, influencing everything from enzyme kinetics and fidelity to the specificity of the amplification product. This guide objectively compares the performance of PCR across a spectrum of magnesium chloride (MgCl₂) concentrations, synthesizing quantitative data and experimental methodologies to provide researchers with evidence-based optimization strategies. The evaluation is contextualized within the broader thesis of PCR fidelity, offering a framework for scientists in drug development and molecular biology to standardize and enhance their amplification protocols.
In the Polymerase Chain Reaction, the DNA polymerase enzyme is responsible for synthesizing new DNA strands. However, this enzyme does not operate alone; it requires a divalent metal ion cofactor, with Mg²⁺ being the most prevalent and efficient in this role [1]. The Mg²⁺ ion is fundamental to the very mechanism by which nucleotides are stitched together into a polynucleotide chain. Acting at the enzyme's active site, it directly enables the catalysis of the phosphodiester bond—the crucial covalent link between the 3'-hydroxyl group of the growing DNA chain and the 5'-phosphate group of an incoming deoxynucleoside triphosphate (dNTP) [2]. Beyond this primary catalytic role, Mg²⁺ also contributes to the structural integrity of the PCR milieu. It stabilizes the double-stranded structure of DNA and facilitates the specific binding of primers to their complementary template sequences by neutralizing the negative charges on the sugar-phosphate backbones, thereby reducing electrostatic repulsion [1]. Consequently, the concentration of MgCl₂ in a PCR buffer is not a mere ingredient but a central parameter that requires precise optimization to balance reaction efficiency, specificity, and fidelity.
Molecular Mechanism: How Mg²⁺ Catalyzes Bond Formation
The catalysis of the phosphodiester bond formation follows a two-metal-ion mechanism that is conserved across DNA polymerases [3]. This mechanism is elegantly illustrated in the crystal structures of polymerase ternary complexes, where two magnesium ions are positioned in the active site, coordinated by invariant aspartate residues.
The Two-Metal-Ion Mechanism
In the Klenow fragment of DNA polymerase I, for example, the carboxylate ligands Asp705 and Asp882 anchor the two catalytic metal ions [3]. These two ions, termed Metal A and Metal B, play distinct but cooperative roles:
- Metal A (Catalytic Metal): This ion primarily coordinates the 3'-OH group of the primer strand. By binding to the oxygen atom, it facilitates the deprotonation of the 3'-hydroxyl group and enhances its nucleophilicity, enabling the attack on the α-phosphate of the incoming dNTP [3] [4].
- Metal B (dNTP-Bound Metal): This ion enters the active site complexed with the incoming dNTP. It coordinates the β- and γ-phosphate oxygens of the nucleotide, stabilizing the structure of the triphosphate and assisting in the neutralization of negative charge buildup as the pyrophosphate leaving group departs [3].
This orchestrated action significantly lowers the activation energy required for the nucleophilic substitution reaction, making the formation of the phosphodiester bond kinetically feasible under physiological and PCR conditions. The role of Mg²⁺ is so critical that its removal or replacement with non-catalytic metal ions renders the polymerase enzymatically dead [1].
Pre-Chemistry Steps and Mg²⁺ Requirements
Kinetic studies reveal that the requirement for Mg²⁺ is not uniform across all steps of the polymerase reaction cycle. Research on Pol I(KF) indicates that the early pre-chemistry steps, including a DNA rearrangement and the closing of the fingers subdomain, can proceed at very low Mg²⁺ concentrations [3]. The fingers-closing step, which creates the active-site geometry for catalysis, is dependent on the aspartate residue (Asp882) that later coordinates one of the metal ions. However, the full complement of Mg²⁺, particularly the entry of the second metal ion (Metal A) into the active site, is mandatory for the covalent nucleotide addition step to occur [3].
The following diagram summarizes the two-metal-ion mechanism of phosphodiester bond formation catalyzed by DNA polymerase:
Comparative Performance: PCR Efficiency Across Mg²⁺ Concentrations
The concentration of MgCl₂ in a PCR is a decisive factor that directly influences the outcome of the amplification. A systematic meta-analysis of 61 peer-reviewed studies established a clear quantitative relationship between MgCl₂ concentration and PCR performance, with optimal ranges identified between 1.5 and 3.0 mM [5]. Deviations from this optimal window have predictable and measurable consequences on reaction efficiency and product specificity.
Quantitative Effects of Mg²⁺ Concentration
The table below synthesizes experimental data on how varying MgCl₂ concentrations impact key PCR performance metrics.
Table 1: Impact of Magnesium Chloride Concentration on PCR Performance Metrics
| MgCl₂ Concentration | DNA Polymerase Activity | Primer Annealing & Melting Temperature (Tm) | Reaction Specificity | Expected Gel Electrophoresis Result |
|---|---|---|---|---|
| Low (< 1.5 mM) | Significantly reduced due to insufficient cofactor [1] [6] | Tm decreased; primers fail to bind stably [1] | High, but yield is severely compromised | Weak amplification, smearing, or no visible product [7] |
| Optimal (1.5 - 3.0 mM) | Maximal catalytic activity; efficient dNTP incorporation [5] | Tm stabilized; specific primer-template binding facilitated [1] [5] | High; specific target amplification dominates | A single, sharp band of the expected size [1] |
| High (> 3.0 - 4.5 mM) | High, but error-prone; may incorporate mismatched nucleotides | Tm increased; promotes non-specific primer binding [1] [6] | Low; non-specific amplification and primer-dimer formation | Multiple bands, non-specific products, or primer dimers [1] [6] |
The meta-analysis revealed a logarithmic relationship between MgCl₂ concentration and DNA melting temperature, with every 0.5 mM increase within the optimal range associated with a 1.2 °C increase in Tm [5]. This quantitatively underscores the ion's role in stabilizing nucleic acid duplexes.
Template-Dependent Optimization
It is crucial to recognize that the "optimal" concentration is not universal but must be tailored to the specific reaction conditions. A key factor is the complexity of the DNA template. The same meta-analysis found that genomic DNA templates, being more complex, generally require higher Mg²⁺ concentrations than simpler templates like plasmid DNA or cDNA [5]. Furthermore, challenging scenarios such as amplifying templates with high GC content, using suboptimal primers, or dealing with DNA extracts containing PCR inhibitors (which can chelate and reduce the availability of free Mg²⁺) often necessitate empirical optimization and potentially the use of concentrations higher than the standard 2 mM [1].
Experimental Protocols for Mg²⁺ Optimization
A systematic approach to Mg²⁺ optimization is fundamental for establishing robust and reproducible PCR protocols, especially within a research thesis framework focused on PCR fidelity.
Standard Mg²⁺ Titration Experiment
This protocol provides a detailed methodology for determining the optimal MgCl₂ concentration for a specific PCR assay.
Objective: To empirically determine the MgCl₂ concentration that yields the highest specificity and yield for a given primer-template system.
Research Reagent Solutions: Table 2: Essential Reagents for Mg²⁺ Optimization Experiments
| Reagent | Function in the Experiment | Typical Stock Concentration |
|---|---|---|
| 10X PCR Buffer (without MgCl₂) | Provides the core reaction environment (pH, salts). Using a Mg-free buffer is essential for a controlled titration. | - |
| MgCl₂ Solution | The variable being tested; source of the Mg²⁺ cofactor. | 25 mM or 50 mM |
| DNA Polymerase | Catalyzes DNA synthesis; its activity is directly dependent on Mg²⁺. | 1-5 U/μL |
| dNTP Mix | Building blocks for new DNA strands; equimolar amounts are critical. | 10 mM each |
| Forward & Reverse Primers | Define the start and end of the target amplicon. | 10 μM |
| DNA Template | The sample containing the target sequence to be amplified. | Varies (e.g., 10-100 ng/μL) |
| Nuclease-Free Water | Solvent to bring the reaction to the final volume. | - |
Methodology:
- Reaction Setup: Prepare a master mix containing all PCR components except the DNA template and MgCl₂. This includes nuclease-free water, 10X Mg-free PCR buffer, dNTP mix, primers, and DNA polymerase. Aliquoting the master mix ensures consistency across reactions.
- Mg²⁺ Titration: Dispense equal volumes of the master mix into individual PCR tubes. Add a varying volume of MgCl₂ stock solution to each tube to create a concentration gradient. A standard range is from 0.5 mM to 5.0 mM in 0.5 mM increments. For a 50 μL reaction with a 25 mM MgCl₂ stock, this would involve adding 1 μL to achieve 0.5 mM, 2 μL for 1.0 mM, and so on, adjusting the water volume accordingly.
- Template Addition: Add the DNA template to each tube. Include a negative control (no template DNA) for one of the Mg²⁺ concentrations to check for contamination.
- Thermal Cycling: Run the PCR using the predetermined cycling parameters (denaturation, annealing, and extension temperatures and times).
- Product Analysis: Analyze the PCR products using agarose gel electrophoresis. Visualize the DNA bands under UV light after staining with an intercalating dye.
Data Analysis: Identify the MgCl₂ concentration that produces a single, intense band of the expected amplicon size with minimal to no background smearing or non-specific bands. This concentration is considered optimal for that specific assay.
Advanced Kinetic Analysis of Pre-Chemistry Steps
For a deeper investigation into the role of Mg²⁺ in the polymerization mechanism itself, as discussed in the context of PCR fidelity, more advanced biophysical assays can be employed.
Objective: To dissect the effect of Mg²⁺ on the individual pre-chemistry conformational changes of the DNA polymerase.
Methodology (based on [3]): This protocol utilizes stopped-flow fluorescence assays with specifically labeled DNA or enzyme.
- Fluorescent Reporter Systems:
- 2-Aminopurine (2-AP) Assay: A 2-AP base is incorporated into the DNA template near the primer-template junction. Its fluorescence is quenched when in a hydrophobic environment (e.g., within the polymerase active site). A fluorescence change reports on the early DNA rearrangement step (step 2.1 in the Pol I(KF) pathway).
- FRET-based Assay: The DNA polymerase is site-specifically labeled with a fluorophore (e.g., IAEDANS) on the fingers subdomain. A change in fluorescence resonance energy transfer (FRET) efficiency reports on the open-to-closed fingers subdomain transition (step 2.2).
- Kinetic Measurements: The polymerase and DNA are rapidly mixed with dNTPs and Mg²⁺ in a stopped-flow instrument, and the fluorescence change is monitored over time. These experiments are repeated at various, precisely controlled low concentrations of MgCl₂ or MnCl₂.
- Data Interpretation: As demonstrated in [3], these assays can determine which pre-chemistry steps (DNA rearrangement, fingers-closing) can proceed at low Mg²⁺ concentrations and which absolutely require the full catalytic complement of ions.
The Scientist's Toolkit: Research Reagent Solutions
Successful investigation into Mg²⁺ effects requires high-quality, specific reagents. The following table details essential materials.
Table 3: Key Research Reagent Solutions for Investigating Mg²⁺ in PCR
| Category | Specific Product Examples | Critical Function & Rationale |
|---|---|---|
| Specialized Buffers | Mg-free 10X PCR Buffer, Optimization Kits | Provides a consistent baseline (pH, KCl, Tris-HCl) without the variable of Mg²⁺, enabling precise titration. |
| Magnesium Solutions | MgCl₂ (25/50 mM), MgSO₄ (for some polymerases) | The independent variable; a high-purity, nuclease-free stock solution is mandatory for accurate results. |
| DNA Polymerases | Standard Taq, High-Fidelity/Proofreading, Hot-Start | Different enzymes may have varying affinities for Mg²⁺. Hot-start polymerases are particularly useful for increasing specificity during optimization. |
| Nucleotides | PCR-grade dNTP Set (dATP, dCTP, dGTP, dTTP) | High-quality, equimolar dNTPs are essential as they chelate Mg²⁺; inconsistent quality can invalidate results. |
| Analysis Tools | DNA Gel Electrophoresis System, DNA Ladder, Fluorescent Dyes | For visualizing and interpreting the results of the optimization experiment (specific vs. non-specific amplification). |
The role of Mg²⁺ in PCR extends far beyond a simple buffer component; it is the central cofactor without which the fundamental reaction of phosphodiester bond formation cannot proceed. The experimental data clearly demonstrates that a narrow concentration window exists for optimal PCR performance, balancing the imperative of maximal polymerase activity with the necessity for high amplification specificity. Titrating MgCl₂ is therefore not an optional step but a core requirement for developing any rigorous, reliable PCR-based assay.
For researchers and drug development professionals, a deep understanding of this mechanism is critical. In diagnostic applications, inconsistent Mg²⁺ levels can lead to false negatives or positives, compromising result integrity. In cloning and sequencing workflows, suboptimal Mg²⁺ can introduce mutations or generate spurious products, wasting valuable time and resources. Therefore, the systematic optimization of Mg²⁺ concentration, as outlined in this guide, should be considered a foundational practice in any molecular biology laboratory, directly contributing to the reliability and fidelity of scientific data.
Magnesium ions (Mg²⁺) serve as indispensable cofactors in polymerase chain reaction (PCR) and related molecular biology techniques, fundamentally influencing both the structural dynamics of nucleic acids and the catalytic efficiency of processing enzymes. The divalent nature of Mg²⁺ allows it to act as a electrostatic bridge between negatively charged phosphate groups on DNA and amino acid residues within enzyme active sites. In PCR specifically, Mg²⁺ is supplied in the form of magnesium chloride (MgCl₂) and plays a dual role: facilitating the enzymatic activity of DNA polymerase and stabilizing the hybrid formed between primers and their template DNA sequences [1] [8]. The concentration of Mg²⁺ becomes a critical experimental variable, with significant implications for the specificity, yield, and fidelity of DNA amplification [9].
The mechanistic basis of Mg²⁺ function extends beyond simple charge neutralization. Current research explores how Mg²⁺ participates in the precise molecular choreography of DNA synthesis, from initial primer binding to the final chemical step of nucleotide incorporation. Understanding these biochemical mechanisms provides researchers with a rational framework for optimizing reaction conditions, particularly when aiming to distinguish between closely related sequences or when working with challenging templates such as those with high GC content. This guide systematically compares the effects of varying Mg²⁺ concentrations on PCR performance, providing experimental data and methodologies relevant to research scientists and drug development professionals evaluating PCR fidelity.
Molecular Mechanisms of Mg²⁺ Action
Stabilization of Primer-Template Hybrids
The hybridization between a primer and its complementary template DNA is a fundamental step in PCR that depends critically on Mg²⁺ concentration. Primers are short, single-stranded DNA oligonucleotides that must specifically bind to their target sequences before DNA polymerase can initiate synthesis. The backbone of DNA and RNA is highly negatively charged due to phosphate groups, creating electrostatic repulsion between the primer and template strands that must be overcome for stable duplex formation [1].
Mg²⁺ addresses this challenge through its positive charge, which neutralizes the electrostatic repulsion between the negatively charged phosphate groups of the primer and template DNA strands [1]. This shielding effect significantly increases the melting temperature (Tm) of the primer-template duplex, defined as the temperature at which half of the DNA duplexes dissociate into single strands [1]. By increasing Tm, Mg²⁺ promotes more stable binding between primers and their intended targets, particularly during the annealing phase of PCR when temperatures are lowered to facilitate these interactions.
The specificity of primer binding is exquisitely sensitive to Mg²⁺ concentration. At optimal concentrations, Mg²⁺ promotes specific primer-template interactions while maintaining sufficient stringency to prevent non-specific binding. However, when Mg²⁺ concentration becomes excessive, the stabilization becomes non-discriminative, allowing primers to bind to partially complementary sequences with reduced fidelity [9]. This loss of specificity manifests experimentally as multiple bands or smeared products in gel electrophoresis, complicating result interpretation and potentially leading to false conclusions in diagnostic applications [9] [7].
Activation of DNA Polymerase Enzymes
The catalytic activity of DNA polymerases, including the commonly used Taq DNA polymerase, depends fundamentally on Mg²⁺ ions that participate directly in the enzymatic mechanism of nucleotide incorporation. Structural and kinetic studies have revealed that DNA polymerases typically employ a two-metal-ion mechanism to facilitate the nucleotidyl transfer reaction [10].
In this conserved mechanism, one metal ion (Mg·dNTP) forms a tight complex with the incoming deoxynucleoside triphosphate (dNTP) by coordinating with non-bridging oxygens across all three phosphate groups [10]. A second catalytic metal ion (Mg²⁺) plays a distinct role by reducing the pKa of the 3'-OH group of the primer terminus, thereby activating the oxygen nucleophile for attack on the α-phosphate of the incoming nucleotide [10]. The coordinated action of these two metal ions, along with water molecules and acidic residues in the active site, stabilizes the pentacovalent transition state during phosphodiester bond formation [10].
Kinetic analysis of HIV reverse transcriptase has demonstrated that the Mg·dNTP complex is necessary and sufficient to induce the conformational change from an open to closed enzyme state, while the second catalytic Mg²⁺ binds subsequently to stabilize the closed state and stimulate the chemical reaction [10]. This weak binding of the catalytic Mg²⁺ contributes to fidelity by allowing sampling of correctly aligned substrates without perturbing the equilibrium for nucleotide binding at physiological Mg²⁺ concentrations [10]. The concentration of free Mg²⁺ significantly impacts catalytic efficiency, with increases from 0.25 to 10 mM shown to enhance nucleotide specificity (kcat/Km) by 12-fold, largely by increasing the rate of chemistry relative to nucleotide release [10].
Table 1: Comparative Effects of Mg²⁺ Concentration on PCR Performance
| Mg²⁺ Concentration | Specificity | Yield | Common Artifacts | Recommended Applications |
|---|---|---|---|---|
| Low (0.5-1 mM) | High | Low to none | Smearing; weak or no amplification | High-specificity applications with simple templates |
| Optimal (1.5-2.5 mM) | High | High | Sharp, specific bands | Routine PCR, diagnostic applications, qPCR |
| High (3-5+ mM) | Reduced | Variable (often high) | Multiple bands; primer-dimers | Challenging templates (high GC%); may require extensive optimization |
Comparative Experimental Data on Mg²⁺ Effects
Quantitative Analysis of Mg²⁺ Titration
Methodical titration of Mg²⁺ concentration represents a fundamental optimization step in PCR protocol development, with measurable effects on amplification efficiency and specificity. Experimental data demonstrates that the optimal Mg²⁺ concentration varies significantly depending on template composition, primer characteristics, and the specific DNA polymerase employed [1] [9].
For standard PCR reactions, MgCl₂ concentrations typically range from 1.0 to 5.0 mM, with 2.0 mM being the most commonly employed starting point for optimization [1]. However, specific applications may require deviation from this standard. Templates with high GC content often benefit from increased Mg²⁺ concentrations (up to 3-4 mM) to overcome secondary structures that impede amplification [1]. Similarly, reactions utilizing suboptimal primers or those containing PCR inhibitors may require elevated Mg²⁺ to compensate for reduced enzyme efficiency or Mg²⁺ sequestration by contaminants [1].
Quantitative analysis reveals that fine-tuning Mg²⁺ concentration can improve amplification specificity by over 40% and yield by up to three-fold [8]. These improvements are quantifiable through multiple metrics, including the cycle threshold (Ct) in real-time PCR, endpoint fluorescence intensity, and band clarity in gel electrophoresis. Kinetic models of qPCR further elucidate how Mg²⁺ concentration influences amplification efficiency across cycles, with primer concentration playing a complementary role in determining reaction progress [11].
Table 2: Mg²⁺ Concentration Effects on DNA Polymerase Kinetics
| Kinetic Parameter | Effect of Low Mg²⁺ | Effect of High Mg²⁺ | Mechanistic Basis |
|---|---|---|---|
| Catalytic Rate (kcat) | Reduced | Increased, but with reduced fidelity | Altered chemistry rate and nucleotide alignment |
| Apparent Km | Increased | Decreased | Enhanced dNTP binding but reduced specificity |
| Processivity | Reduced | Moderately increased | Improved primer-template stability |
| Error Rate | Variable | Significantly increased | Reduced discrimination against mismatched nucleotides |
Effects on PCR Fidelity and Specificity
The fidelity of DNA amplification—defined as the accuracy with which DNA polymerase incorporates complementary nucleotides—shows a strong dependence on Mg²⁺ concentration. While Mg²⁺ is essential for the catalytic function of DNA polymerases, improper concentrations can dramatically increase error rates and promote non-specific amplification [9].
At elevated Mg²⁺ concentrations (typically >3-4 mM), the stabilization of primer-template interactions becomes less discriminative, allowing primers to bind to sequences with partial complementarity [9]. This non-specific binding results in amplification of unintended products, which compete with target amplification and reduce the yield and purity of the desired product [9]. Experimental evidence demonstrates that excessive Mg²⁺ promotes the formation of primer-dimers and other amplification artifacts, observable as multiple bands or smeared backgrounds in agarose gel electrophoresis [9] [7].
Conversely, insufficient Mg²⁺ concentrations (<1.0 mM) impair the catalytic activity of DNA polymerase, leading to reduced processivity and incomplete extension products [1] [7]. These truncated amplification products manifest as smearing in gel electrophoresis, beginning at the expected product size and extending downward, rather than as discrete bands [7]. In severe cases, Mg²⁺ deficiency can completely prevent amplification, yielding no detectable product [1].
Recent advances in enzyme engineering have produced novel DNA polymerase variants with enhanced capabilities, such as the reverse transcriptase activity demonstrated by certain engineered Taq polymerase variants [12]. These specialized enzymes may exhibit distinct Mg²⁺ optima that differ from conventional polymerases, necessitating re-optimization of reaction conditions when implementing new enzyme systems [12].
Experimental Protocols for Mg²⁺ Optimization
Standard Mg²⁺ Titration Methodology
A systematic approach to Mg²⁺ titration provides researchers with a reliable method for establishing optimal PCR conditions for novel primer-template systems or when working with unfamiliar DNA polymerases. The following protocol outlines a standardized procedure for Mg²⁺ optimization:
Reagents and Equipment:
- 10X PCR Buffer (without MgCl₂)
- 25 mM MgCl₂ stock solution
- DNA template (diluted to appropriate concentration)
- Forward and reverse primers (10 μM each)
- dNTP mix (10 mM total)
- DNA polymerase (concentration per manufacturer's recommendation)
- Sterile nuclease-free water
- PCR thermal cycler
- Agarose gel electrophoresis system
Procedure:
- Prepare a master mix containing all PCR components except MgCl₂ and DNA template. Calculate volumes for n+1 reactions to account for pipetting error.
- Aliquot the master mix into 8 PCR tubes labeled with intended Mg²⁺ concentrations (0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, and 4.0 mM final concentration).
- Add the appropriate volume of 25 mM MgCl₂ stock to each tube to achieve the target concentrations.
- Add DNA template to each tube and mix thoroughly by gentle pipetting.
- Conduct PCR amplification using cycling parameters appropriate for the primer-template system.
- Analyze results by agarose gel electrophoresis, comparing band intensity, specificity, and absence of secondary products across the concentration gradient.
- Select the Mg²⁺ concentration that produces the strongest specific amplification with minimal background or non-specific products.
This protocol can be adapted for quantitative PCR applications by substituting gel analysis with fluorescence monitoring and calculating reaction efficiency based on standard curves or comparative Ct methods.
Advanced Kinetic Analysis of Mg²⁺-Dependent Amplification
For research applications requiring precise quantification of amplification efficiency, kinetic modeling approaches provide sophisticated tools for analyzing Mg²⁺ effects throughout the PCR process rather than solely at endpoint. The following methodology adapts established kinetic models for studying Mg²⁺-dependent amplification [11]:
Theoretical Framework: The stepwise kinetic equilibrium model treats each PCR cycle as a series of equilibrium reactions, with efficiency at cycle n (Eₙ) dependent on reactant concentrations. The model incorporates primer concentration (P) and Mg²⁺ concentration as variables affecting annealing efficiency. The target concentration at cycle n is given by:
Tₙ = T₀ × Π(1 + Eᵢ) from i=1 to n
Where Eᵢ represents the cycle-dependent efficiency influenced by Mg²⁺ concentration through its effects on primer-template stability and polymerase activity.
Experimental Design:
- Perform qPCR reactions with identical primer and template concentrations across a Mg²⁺ concentration gradient (0.5-4.0 mM in 0.5 mM increments).
- Use intercalating dyes (SYBR Green) rather than hydrolysis probes to directly monitor double-stranded DNA accumulation.
- Collect fluorescence data throughout all amplification cycles with high data density.
- Apply kinetic modeling to calculate efficiency values for each cycle at different Mg²⁺ concentrations.
- Compare efficiency curves to identify Mg²⁺ concentrations that maintain high efficiency through the maximum number of cycles before plateau.
This approach provides a more comprehensive understanding of how Mg²⁺ concentration influences not just endpoint yield but the kinetic progression of amplification, enabling finer optimization for demanding applications such as rare allele detection or absolute quantification.
Research Reagent Solutions
Table 3: Essential Reagents for Investigating Mg²⁺ Effects in PCR
| Reagent | Function | Considerations for Mg²⁺ Studies |
|---|---|---|
| MgCl₂ (25-100 mM stock solutions) | Source of Mg²⁺ ions | Use high-purity, nuclease-free preparations; concentration must be verified for accurate titration |
| Mg²⁺-free PCR buffers | Reaction environment without Mg²⁺ | Essential for controlled studies; often supplied by polymerase manufacturers |
| dNTP mix | Nucleotide substrates | Compete with primers for Mg²⁺ binding; maintain constant dNTP concentration during Mg²⁺ titration |
| DNA polymerases | Catalytic enzyme | Different polymerases have distinct Mg²⁺ optima; note proofreading activity may alter requirements |
| Control DNA templates | Amplification substrate | Use well-characterized templates with known amplification characteristics |
| Gradient PCR instrument | Thermal cycler | Enables simultaneous testing of multiple annealing temperatures alongside Mg²⁺ optimization |
| qPCR systems with intercalating dyes | Real-time monitoring | Allows kinetic analysis of amplification efficiency at different Mg²⁺ concentrations |
Mg²⁺ in Emerging Diagnostic Technologies
The critical role of Mg²⁺ in nucleic acid amplification extends beyond conventional PCR to emerging diagnostic technologies that require precise optimization for clinical applications. Engineered DNA polymerase variants with specialized functions, such as the novel Taq polymerase variants capable of both reverse transcription and DNA amplification, demonstrate distinct Mg²⁺ requirements that must be characterized during assay development [12]. These single-enzyme systems eliminate the need for viral reverse transcriptases in RT-PCR applications but may necessitate re-optimization of Mg²⁺ concentrations to balance dual enzymatic activities [12].
CRISPR-based diagnostic systems (CRISPRdx) represent another technological advancement where Mg²⁺ plays a crucial role, particularly in reactions combining isothermal amplification with CRISPR-mediated detection [13]. While CRISPR systems offer exceptional potential for point-of-care testing with single-nucleotide resolution, their performance depends on appropriate Mg²⁺ concentrations that support both amplification and Cas enzyme activity [13]. The development of multiplex detection systems, capable of simultaneously identifying multiple RNA targets with a single enzyme, further underscores the importance of comprehensive reaction optimization including Mg²⁺ titration [12].
Diagram 1: Molecular Interactions of Mg²⁺ in DNA Synthesis. This diagram illustrates the dual roles of Mg²⁺ in stabilizing primer-template hybrids and participating in the two-metal-ion catalytic mechanism of DNA polymerases.
The biochemical mechanisms through which Mg²⁺ stabilizes primer-template hybrids and modulates enzyme kinetics represent fundamental aspects of nucleic acid amplification technology with direct implications for research and diagnostic applications. The experimental data and methodologies presented in this comparison guide demonstrate that Mg²⁺ concentration systematically influences PCR specificity, efficiency, and fidelity through defined molecular interactions. Optimal Mg²⁺ concentrations balance the competing demands of primer-template stabilization and enzymatic activity, with significant deviations in either direction producing characteristic artifacts that compromise experimental results.
The continued development of novel nucleic acid amplification systems, including engineered polymerases with dual functionality and CRISPR-based detection platforms, maintains the importance of thorough Mg²⁺ optimization as an essential step in assay development. Researchers pursuing PCR fidelity studies between different magnesium concentrations should employ the systematic titration approaches and kinetic analyses outlined in this guide to establish rigorous, reproducible reaction conditions suitable for their specific applications. Through methodical optimization and understanding of the underlying biochemical principles, scientists can harness the full potential of Mg²⁺-dependent amplification while minimizing artifacts and maintaining the precision required for advanced molecular diagnostics and research applications.
In molecular biology and genetic research, the accuracy of polymerase chain reaction (PCR) is paramount. PCR fidelity refers to the ability of a DNA polymerase to accurately replicate a DNA template without introducing errors during amplification. This characteristic is quantified as an error rate, representing the frequency of misincorporated nucleotides per base per doubling event. For applications ranging from cloning and next-generation sequencing to diagnostic assay development and functional gene analysis, high-fidelity PCR is essential to ensure that amplified products truly represent the original template sequence. This guide objectively compares the performance of various DNA polymerases, examining the experimental data that underpins our understanding of PCR fidelity, with particular attention to how factors like magnesium concentration influence error rates.
Measuring PCR Fidelity: Methodologies and Experimental Data
The assessment of DNA polymerase fidelity relies on several established methodologies, each with distinct protocols, advantages, and limitations. Understanding these methods is critical for interpreting comparative fidelity data.
Established Fidelity Assay Protocols
1. LacZα Complementation Assay (Blue/White Screening) This method utilizes a PCR-amplified fragment of the lacZα gene, which is then cloned into a vector and transformed into bacteria [14] [15].
- Workflow: Amplify the lacZα gene → Clone into vector → Transform into E. coli → Plate on X-Gal indicator plates → Score blue (error-free) and white (mutated) colonies [14].
- Data Analysis: The mutation frequency is calculated based on the ratio of white to total colonies. The error rate is then determined after accounting for the number of effective amplification cycles [14].
- Limitations: The assay is biased, as only mutations within a specific 349-base region of the 1.9 kb lacZ gene result in a color change, leaving many errors undetected [14] [15].
2. Sanger Sequencing of Cloned PCR Products This approach involves sequencing individual cloned PCR products to identify all mutations within the amplified sequence [14] [15].
- Workflow: Amplify a target gene → Clone PCR products → Pick individual colonies → Prepare plasmid DNA → Sequence using Sanger method [15].
- Data Analysis: The total number of mutations is counted across the total number of bases sequenced. The error rate is calculated considering the number of doublings during PCR [14] [15].
- Limitations: Lower throughput and higher cost per base compared to next-generation sequencing methods, making it less suitable for measuring the very low error rates of high-fidelity enzymes with statistical confidence [14].
3. Next-Generation Sequencing (NGS) Assays NGS platforms, such as Illumina and Pacific Biosciences (PacBio), enable deep sequencing of PCR amplicons without a cloning step, providing millions of reads for robust statistical analysis [14] [16].
- PacBio SMRT Sequencing Workflow: Perform polymerase primer extension → Prepare library without PCR amplification → Perform Single Molecule, Real-Time (SMRT) sequencing → Generate circular consensus sequencing (CCS) reads for high accuracy → Align reads to a reference sequence to identify errors [14] [16].
- Data Analysis: Errors are identified by comparing each highly accurate consensus read to the known reference sequence. The background error rate for this method is extremely low (~9.6 × 10⁻⁸), making it suitable for quantifying ultra-high-fidelity polymerases [14].
- Advantages: This high-throughput method provides a comprehensive error profile (substitutions, indels) and can directly sequence PCR products, avoiding biases introduced by bacterial cloning [14] [16].
Comparative Fidelity Data of DNA Polymerases
Experimental data from the methodologies above reveal significant differences in performance between common DNA polymerases. The following table consolidates quantitative findings from PacBio SMRT sequencing, which offers one of the most accurate comparisons available [14].
Table 1: DNA Polymerase Fidelity Measured by PacBio SMRT Sequencing
| DNA Polymerase | Substitution Rate (errors/base/doubling) | Accuracy (1/Substitution Rate) | Fidelity Relative to Taq |
|---|---|---|---|
| Q5 High-Fidelity | 5.3 × 10⁻⁷ | 1,870,763 | 280X |
| Phusion High-Fidelity | 3.9 × 10⁻⁶ | 255,118 | 39X |
| Deep Vent | 4.0 × 10⁻⁶ | 251,129 | 44X |
| Pfu | 5.1 × 10⁻⁶ | 195,275 | 30X |
| PrimeSTAR GXL | 8.4 × 10⁻⁶ | 118,467 | 18X |
| KOD | 1.2 × 10⁻⁵ | 82,303 | 12X |
| Kapa HiFi HotStart | 1.6 × 10⁻⁵ | 63,323 | 9.4X |
| Taq (reference) | 1.5 × 10⁻⁴ | 6,456 | 1X |
| Deep Vent (exo-) | 5.0 × 10⁻⁴ | 2,020 | 0.3X |
Data adapted from NEB SMRT sequencing assays [14].
The data demonstrates that proofreading polymerases (those with 3'→5' exonuclease activity) like Q5, Phusion, and Pfu significantly outperform non-proofreading enzymes like Taq. The critical role of proofreading is starkly illustrated by the 125-fold increase in the error rate when the proofreading domain of Deep Vent is disabled (Deep Vent exo-) [14].
The Impact of Reaction Conditions on PCR Fidelity
While the choice of polymerase is a primary determinant of PCR accuracy, biochemical conditions within the reaction tube profoundly influence the observed error rate.
The Critical Role of Magnesium Concentration
Magnesium ions (Mg²⁺) are an essential cofactor for DNA polymerase activity, but their concentration must be carefully optimized. Research on reverse transcriptase fidelity provides a clear paradigm for how Mg²⁺ affects accuracy [17].
- Physiological vs. Standard Conditions: The fidelity of HIV-1 reverse transcriptase (HIV RT) is typically severalfold lower in standard in vitro assays (using 5-10 mM Mg²⁺) than in cellular infection models. This discrepancy has been reconciled by testing fidelity at a more physiological free Mg²⁺ concentration for lymphocytes (~0.25 mM) [17].
- Experimental Findings: Under high-Mg²⁺ conditions (6 mM), HIV RT fidelity was low. However, when assayed at 0.25 mM Mg²⁺, its error rate decreased by 5 to 10 times, making it comparable to the fidelity of Moloney murine leukemia virus (MuLV) RT, which was itself insensitive to Mg²⁺ concentration [17]. This indicates that the "low fidelity" reputation of HIV RT is an artifact of non-physiological assay conditions and that Mg²⁺ is a key modulator of accuracy.
- Mechanism: Mg²⁺ is a direct coordinator of the nucleotidyl transfer reaction in the polymerase active site. Deviations from the optimal concentration can disrupt the delicate balance of reaction kinetics, potentially increasing misincorporation and decreasing the enzyme's ability to discriminate against incorrect nucleotides.
Other Influential Factors
- dNTP Concentration: Imbalanced or excessively high dNTP pools can force polymerases to incorporate incorrect nucleotides more readily [17] [18].
- Buffer pH and Salt Conditions: The ionic strength and pH of the reaction buffer can affect the stability of primer-template binding and the processivity of the enzyme [18].
- Thermal Cycling Parameters: The duration of the extension step must be sufficient for complete synthesis, but excessively long times may increase opportunities for misincorporation [18].
Visualizing the PCR Fidelity Workflow
The following diagram illustrates the integrated workflow for assessing polymerase fidelity, from biochemical reaction to sequencing-based analysis.
A Researcher's Toolkit for Fidelity Studies
Table 2: Essential Research Reagents and Materials
| Item | Function in Fidelity Assays |
|---|---|
| High-Fidelity DNA Polymerase | Engineered enzymes with high intrinsic accuracy and proofreading activity (e.g., Q5, Pfu) to minimize background errors during amplicon generation [14] [19]. |
| Control DNA Polymerase | Enzymes with known, lower fidelity (e.g., Taq) to serve as a benchmark for comparison in fidelity studies [14] [15]. |
| Target DNA Template | A well-characterized, error-free plasmid (e.g., containing lacZ or another target gene) used as a substrate for amplification to track newly introduced mutations [14] [15]. |
| dNTPs | Deoxynucleoside triphosphates, the building blocks for DNA synthesis; their quality and concentration must be controlled to prevent induced errors [17] [18]. |
| MgCl₂ Solution | A critical cofactor for polymerase activity; concentration must be carefully titrated and optimized, as it is a major variable affecting fidelity [17] [18]. |
| Cloning Kit | For LacZ and Sanger sequencing assays, kits are needed to clone PCR products into a vector for subsequent transformation and analysis [14] [15]. |
| Sequencing Platform | Access to Sanger or Next-Generation Sequencing (e.g., PacBio SMRT) services or instrumentation is required for definitive error detection and quantification [14] [16]. |
Defining PCR fidelity requires a comprehensive understanding of error rates, the mechanisms that underpin them, and the experimental methods used for quantification. Data unequivocally shows that proofreading DNA polymerases like Q5 and Phusion can offer a 280-fold and 39-fold increase in accuracy, respectively, over standard Taq polymerase [14]. However, the innate fidelity of an enzyme is not the sole determinant of success in the lab. Reaction conditions, particularly magnesium concentration, are powerful modulators of error rates, as demonstrated by the significant fidelity improvements in HIV RT under physiological Mg²⁺ levels [17]. For researchers in drug development and diagnostics, where precision is critical, selecting a high-fidelity polymerase and meticulously optimizing the reaction environment are both essential steps to ensure the integrity of amplified DNA and the validity of downstream results.
In polymerase chain reaction (PCR), magnesium chloride (MgCl₂) serves not merely as a reaction component but as a fundamental regulator of enzymatic fidelity, directly determining the balance between amplification success and failure. As an essential cofactor for thermostable DNA polymerases, Mg²⁺ ions influence reaction kinetics at multiple levels, from primer-template binding to nucleotide incorporation efficiency [8]. The concentration of this divalent cation represents one of the most crucial variables in PCR optimization, governing the thermodynamic balance between specificity, yield, and accuracy [5] [20]. Understanding its dual nature—where insufficient Mg²⁺ limits polymerase activity while excess promotes non-specific amplification—is essential for researchers demanding reproducible, high-quality results in molecular diagnostics, gene cloning, and drug development applications [18].
This guide examines the quantitative relationships between Mg²⁺ concentration and key PCR performance metrics, drawing upon recent meta-analyses and comparative studies to provide evidence-based optimization strategies. We present structured experimental data and methodologies to empower research scientists in making informed decisions when designing amplification protocols across various applications and template types.
Quantitative Effects of Mg²⁺ on PCR Performance Parameters
Mg²⁺ Concentration Relationships with Specificity and Yield
The relationship between MgCl₂ concentration and PCR outcomes follows a definable optimum that varies according to template characteristics and reaction composition. Recent meta-analyses of 61 peer-reviewed studies established clear quantitative relationships that can guide initial optimization efforts [5].
Table 1: Optimal MgCl₂ Concentration Ranges for Different Template Types
| Template Type | Optimal MgCl₂ Range (mM) | Key Considerations | Effect of 0.5 mM Increase |
|---|---|---|---|
| Genomic DNA | 2.0 - 3.0 mM | Higher complexity requires increased Mg²⁺ | +1.2°C Tm increase [5] |
| Plasmid DNA | 1.5 - 2.5 mM | Lower complexity enables reduced Mg²⁺ | +1.2°C Tm increase [5] |
| GC-Rich Templates | 2.5 - 3.5 mM | Enhanced stability demands higher Mg²⁺ | Greater stabilization effect [20] |
| Standard Amplicons | 1.5 - 2.0 mM | 100-500 bp fragments | +1.2°C Tm increase [5] [18] |
Mg²⁺ concentration directly influences DNA melting temperature (Tm) through a demonstrated logarithmic relationship. Within the optimal range of 1.5-3.0 mM, each 0.5 mM increment in MgCl₂ concentration produces a consistent 1.2°C increase in melting temperature [5]. This property becomes particularly critical when amplifying GC-rich sequences or complex genomic DNA, where template secondary structure and stability vary significantly [20].
The duality of Mg²⁺ effects manifests clearly in deviation from optimal concentrations. Insufficient Mg²⁺ (below 1.0 mM) dramatically reduces product yield due to impaired DNA polymerase activity and inefficient primer extension [18] [8]. Conversely, excess Mg²⁺ (above 3.0-4.0 mM) promotes non-specific amplification through reduced primer-stringency and stabilization of mismatched primer-template complexes [5] [18]. This often manifests as multiple bands on electrophoresis gels or smeared backgrounds, indicating amplified off-target products [18].
Magnesium-Dependent Fidelity Variations Across DNA Polymerases
The fidelity of DNA synthesis varies significantly among different polymerase classes, with Mg²⁺ concentration playing a modifying role in error frequency. Comparative studies measuring error rates through direct sequencing of cloned PCR products have established clear fidelity rankings [15].
Table 2: DNA Polymerase Fidelity Comparison Under Optimal Mg²⁺ Conditions
| DNA Polymerase | Error Rate (errors/bp/duplication) | Fidelity Relative to Taq | Recommended [Mg²⁺] Range |
|---|---|---|---|
| Taq | 3.0-5.6 × 10⁻⁵ | 1x | 1.5-2.5 mM [8] |
| AccuPrime-Taq HF | ~1.0 × 10⁻⁵ | ~5x better | Vendor-specific [15] |
| KOD Hot Start | ~1.0 × 10⁻⁶ | ~50x better | Vendor-specific [15] |
| Pfu | 1.0-2.0 × 10⁻⁶ | ~10x better | 2.0-3.0 mM [15] |
| Phusion Hot Start | 4.0-9.5 × 10⁻⁷ | >50x better | Vendor-specific [15] |
High-fidelity enzymes such as Pfu and Phusion exhibit significantly lower error rates than Taq polymerase, attributed to their 3'→5' exonuclease proofreading activity [15]. These polymerases demonstrate particular sensitivity to Mg²⁺ concentration, with narrow optimal ranges that must be maintained to preserve their fidelity advantages. For cloning applications and large-scale projects where sequence accuracy is paramount, selection of proofreading enzymes with precisely optimized Mg²⁺ concentrations becomes essential [15].
Experimental Approaches for Mg²⁺ Optimization
Standardized Titration Methodology
Systematic Mg²⁺ optimization requires a structured titration approach to identify the concentration that balances yield, specificity, and accuracy for specific template-primer-enzyme combinations [18]. The following protocol adapts established methodologies from recent plant pathogen detection studies [21] [22]:
Reaction Setup:
- Prepare a master mix containing all PCR components except MgCl₂
- Aliquot equal volumes into 8 PCR tubes
- Add MgCl₂ to create a concentration gradient (e.g., 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0 mM)
- Include both positive and negative controls
- Perform amplification using standardized cycling conditions
- Analyze products via agarose gel electrophoresis and quantify yield
Assessment Criteria:
- Optimal Specificity: Single, intense band of expected size
- Maximum Yield: Highest product quantity without non-specific amplification
- Accuracy Validation: Direct sequencing of amplified products to verify sequence integrity [15]
This methodological framework was successfully applied in developing detection assays for Fusarium tricinctum, where Mg²⁺ optimization proved critical for achieving sensitive and specific amplification across LAMP, nested PCR, and qPCR platforms [21].
Advanced Workflow for Complex Templates
Challenging templates, including those with high GC content, secondary structure, or low complexity, often require extended optimization strategies [20]. Building upon the standardized titration, additional parameters may require simultaneous optimization:
Enhanced Protocol for Difficult Amplicons:
- Perform initial Mg²⁺ titration (1.0-4.0 mM in 0.5 mM increments)
- Combine with annealing temperature gradient (±5-10°C from calculated Tm)
- Incorporate PCR enhancers (e.g., betaine, DMSO, formamide) at various concentrations
- Evaluate polymerase-specific buffer systems
- Employ touchdown PCR protocols for increased specificity
This comprehensive approach proved essential in recent engineering of novel Taq polymerase variants with reverse transcriptase activity, where precise Mg²⁺ optimization enabled single-enzyme quantitative multiplex RT-PCR without compromising sensitivity or specificity [23].
Diagram: Experimental workflow for systematic optimization of Mg²⁺ concentration in PCR, highlighting iterative assessment of specificity and yield. This structured approach efficiently identifies optimal conditions while controlling for multiple variables.
The Scientist's Toolkit: Essential Reagents for PCR Optimization
Table 3: Key Research Reagent Solutions for Magnesium Optimization Studies
| Reagent/Category | Function in PCR Optimization | Application Notes |
|---|---|---|
| MgCl₂ Stock Solutions (25-100 mM) | Provides Mg²⁺ cofactor for polymerase activity | Use high-purity, nuclease-free; concentration significantly affects fidelity [15] [8] |
| High-Fidelity DNA Polymerases (Pfu, Phusion) | Reduces replication errors through proofreading | Essential for cloning; requires specific Mg²⁺ optimization [15] |
| Standard DNA Polymerases (Taq) | Standard amplification with moderate fidelity | More tolerant of Mg²⁺ variation; lower fidelity [15] |
| dNTP Mix | DNA synthesis substrates | Concentration affects free Mg²⁺ availability; optimize with Mg²⁺ [18] |
| PCR Buffers (with/without Mg²⁺) | Maintains optimal pH and chemical environment | Mg²⁺-free buffers enable precise customization [18] |
| PCR Enhancers (Betaine, DMSO) | Reduces secondary structure, improves efficiency | Particularly valuable for GC-rich templates [20] |
| Quantitative Standards | Enables precise efficiency calculations | Essential for qPCR optimization [24] |
The optimization of Mg²⁺ concentration represents a fundamental compromise between competing PCR performance metrics that must be resolved according to application-specific requirements. Diagnostic applications prioritizing sensitivity may tolerate slightly higher Mg²⁺ concentrations despite potential fidelity costs, while cloning and sequencing applications demand stringent optimization for maximum accuracy [15]. The expanding landscape of specialized polymerase formulations, including novel variants with dual reverse transcriptase and polymerase activities [23], further emphasizes the continuing importance of cation optimization in modern molecular biology.
Future methodological developments will likely continue to refine our understanding of Mg²⁺'s role in PCR fidelity, particularly as automated optimization platforms and machine learning approaches become more widespread. Nevertheless, the principles established through decades of PCR research remain foundational: precise Mg²⁺ control balances the dual nature of this essential cation, enabling researchers to achieve the specificity, yield, and accuracy demanded by contemporary molecular applications.
Reverse transcriptases (RTs) are critical enzymes in molecular biology and virology, playing a pivotal role in retroviral replication and biotechnological applications. For decades, RT studies have been conducted under standardized in vitro conditions optimized for maximum enzymatic activity rather than physiological relevance. This guide objectively compares RT performance under physiological versus standard magnesium (Mg²⁺) conditions, focusing on fidelity implications for PCR, drug development, and virological research. The central thesis demonstrates that physiological Mg²⁺ concentrations profoundly alter RT fidelity and function compared to standard laboratory conditions, with significant implications for experimental interpretation and therapeutic targeting.
Magnesium in Cellular Physiology vs. Laboratory Practice
The Magnesium Discrepancy
The fundamental discrepancy in RT research lies in the concentration of Mg²⁺ used in experimental systems versus actual cellular environments:
- Total Cellular Mg²⁺: Approximately 10 mM or more [25]
- Physiological Free Mg²⁺: ~0.5 mM in lymphocytes and other cell types [25] [26]
- Standard In Vitro Mg²⁺: Typically 5-10 mM for RT assays [25]
This disparity occurs because most intracellular Mg²⁺ is sequestered by nucleotides (particularly ATP, present at 1.3-4.3 mM), nucleic acids, and other complex anions [25] [26]. The critical parameter for enzymatic activity is the concentration of free Mg²⁺ ions, which is substantially lower than the total cellular pool.
Mechanisms of Magnesium in Reverse Transcription
Mg²⁺ serves as an essential cofactor for both catalytic functions of HIV-1 RT:
- Polymerase Activity: Two Mg²⁺ ions separated by ~3.6 Å coordinate substrate binding and catalysis in the polymerase active site, with three aspartate residues (D110, D185, D186) essential for metal ion binding [26].
- RNase H Activity: Divalent metal ions (likely Mg²⁺ in vivo) are essential for RNA template degradation, though the precise number and positioning in the full RT enzyme remains under investigation [26].
The following diagram illustrates the relationship between Mg²⁺ concentrations and their impact on reverse transcription:
Comparative Performance Under Differing Mg²⁺ Conditions
Fidelity and Error Rate Analysis
The most significant difference between physiological and standard Mg²⁺ concentrations concerns RT fidelity. Multiple studies demonstrate that HIV-1 RT exhibits substantially different error rates depending on Mg²⁺ availability:
Table 1: Fidelity Comparison Across Divalent Cations and Concentrations
| Cation | Concentration | Relative Fidelity | Mutation Frequency | Study Type |
|---|---|---|---|---|
| Mg²⁺ | 0.5 mM (Physiological) | Higher | ~4-fold decrease vs. high Mg²⁺ | α-complementation [25] |
| Mg²⁺ | 6 mM (Standard) | Lower | ~4-fold increase vs. low Mg²⁺ | α-complementation [25] |
| Zn²⁺ | 0.4 mM (Optimal) | 2.5× greater than Mg²⁺ | Significant decrease | lacZα-complementation [27] |
| Co²⁺ | 0.25 mM (Optimal) | Similar to Mg²⁺ | No significant change | lacZα-complementation [27] |
| Mn²⁺ | 0.4 mM (Optimal) | Similar to Mg²⁺ | No significant change | lacZα-complementation [27] |
Next-generation sequencing (NGS) analysis using barcoding to determine mutation profiles confirmed these findings, demonstrating an approximately four-fold increase in mutations when HIV-1 RT operated at 6 mM Mg²⁺ compared to 0.5 mM Mg²⁺ on a lacZα template [25]. Unlike α-complementation assays dependent on LacZα activity, the NGS approach scores mutations at all positions and of every type, providing comprehensive mutation spectra.
Enzymatic Efficiency and Processivity
Beyond fidelity, Mg²⁺ concentration significantly impacts overall RT performance:
Table 2: Functional Parameters Affected by Mg²⁺ Concentration
| Parameter | 0.5 mM Mg²⁺ | 6-8 mM Mg²⁺ | Biological Significance |
|---|---|---|---|
| ssDNA Synthesis | More efficient | Less efficient | Impacts completion of reverse transcription [25] [26] |
| RNase H Activity | Indirectly reduced | Enhanced | Affects template stability and premature dissociation [26] |
| NRTI Inhibition | Reduced effectiveness | Optimal incorporation | Influences antiviral drug efficacy [26] |
| Template Degradation | Decreased | Increased | Generates dead-end DNA products at high Mg²⁺ [26] |
At low Mg²⁺ concentrations, reverse transcription of natural templates increases despite dramatically reduced intrinsic polymerase activity under such conditions. This apparent paradox is explained by reduced RNA degradation preventing premature dissociation of template and primer strands that otherwise generate dead-end DNA products [26].
Reverse Transcriptase-Specific Responses
Different RTs exhibit distinct sensitivity to Mg²⁺ concentrations:
- HIV-1 RT (subtype B, A/E, drug-resistant variants): Shows higher fidelity at physiological Mg²⁺ concentrations [25]
- HIV-2 RT: Demonstrates higher fidelity at physiological Mg²⁺ [25]
- Prototype Foamy Virus (PFV) RT: Shows higher fidelity in low Mg²⁺ [25]
- MuLV RT: Displays equivalent fidelity in low and high Mg²⁺ [25]
- AMV RT: Exhibits equivalent fidelity in low and high Mg²⁺ [25]
This variation suggests that MuLV and AMV RTs are outliers, as most tested RTs show greater fidelity in low Mg²⁺ conditions [25].
Experimental Protocols for Mg²⁺ Fidelity Assessment
lacZα-Based α-Complementation Assay
This widely employed method measures mutation rates through functional complementation of the lacZα peptide:
Procedure:
- Template Preparation: Plasmid pBS▽EcoRV567 cleaved with NdeI, with T3 RNA polymerase producing run-off transcripts ~644 nucleotides [25]
- Reaction Conditions: 50 mM Tris-HCl pH 8, 80 mM KCl, 1 mM DTT, 20 μM dNTPs, 0.5 or 6 mM MgCl₂ (final free concentration after correcting for dNTP chelation), 37°C for 45 minutes [25]
- Two-Round DNA Synthesis: First round uses RNA template with 5'-³²P-labelled primer; four identical reactions per condition with 25 nM template and 50 nM primer [25]
- Hybridization: Primer and template hybridized in reaction buffer without dNTPs and MgCl₂ by heating to 65°C for 5 minutes followed by slow cooling [25]
- Mutation Scoring: White or faint blue colonies indicate mutations disrupting LacZα function
Next-Generation Sequencing (NGS) Fidelity Profiling
For comprehensive mutation analysis, NGS approaches provide superior resolution:
Workflow:
- Template Design: lacZα template suitable for RT synthesis
- Barcoding Strategy: Incorporation of molecular barcodes to distinguish true mutations from amplification artifacts
- Library Preparation: Using commercial kits (e.g., NEBNext Ultra II DNA Library Prep Kit for Illumina) with appropriate barcoding oligos [25]
- Sequence Analysis: Alignment to reference sequence with mutation calling at all template positions
This method identified approximately 250,000 mutations across experimental conditions, providing unprecedented resolution of RT error spectra [25].
The following diagram outlines the experimental workflow for assessing Mg²⁺ effects on RT fidelity:
Research Reagent Solutions
Essential materials and reagents for studying Mg²⁺ effects on RT fidelity:
Table 3: Key Research Reagents for Mg²⁺-RT Fidelity Studies
| Reagent/Category | Specific Examples | Function/Application | Experimental Considerations |
|---|---|---|---|
| Reverse Transcriptases | HIV-1 RT (HXB2 strain), HIV-2 RT, PFV RT, MuLV RT, AMV RT | Comparative fidelity studies | Source from commercial suppliers or academic collaborators [25] |
| Template Systems | lacZα-complementation templates, natural RNA templates (HIV-1 MAL genomic RNA) | Fidelity assessment | Homopolymeric templates vs. natural sequences yield different fidelity estimates [25] [26] |
| Nucleotide Analogs | AZTTP, d4TTP, 3TCTP, ddCTP, ddATP | NRTI inhibition studies | Treat with pyrophosphatase to prevent contamination with PPi [26] |
| Mg²⁺ Modulators | ATP, EDTA, EGTA | Manipulating free Mg²⁺ concentrations | ATP binds significant Mg²⁺ pool (physiological relevance) [26] |
| Library Prep Kits | NEBNext Ultra II DNA Library Prep Kit for Illumina | NGS mutation profiling | Includes barcoding oligos for multiplexing [25] |
Implications for Research and Therapeutic Development
Virological and Drug Development Implications
The differential behavior of RT under physiological Mg²⁺ conditions has profound implications:
- Antiretroviral Drug Efficacy: NRTIs show reduced effectiveness at physiological Mg²⁺ concentrations, while NNRTIs demonstrate enhanced activity [25]. This suggests that current in vitro drug screening may misrepresent compound efficacy.
- Cellular Mutation Rates: Discrepancies between in vitro and cellular mutation rates may reflect non-physiological Mg²⁺ concentrations in standard assays [25].
- Viral Evolution: Differences in Mg²⁺ availability between cell types or during the cell cycle might strongly affect HIV-1 replication and mutation rates in vivo [26].
Recommendations for Experimental Design
Based on comparative data, researchers should:
- Utilize Physiological Mg²⁺: Include 0.5-1.0 mM free Mg²⁺ in fidelity assays to better approximate in vivo conditions
- Account for Cation Buffers: Include physiological ATP concentrations (1.3-4.3 mM) that sequester Mg²⁺ and reduce free cation availability [26]
- Validate with Multiple RTs: Test enzyme variants beyond HIV-1 RT, as Mg²⁺ sensitivity varies between retroviral RTs
- Employ Orthogonal Methods: Combine functional assays (α-complementation) with comprehensive sequencing approaches
The discrepancy between physiological and standard in vitro Mg²⁺ conditions represents a critical factor in reverse transcriptase research that has been largely overlooked. Physiological Mg²⁺ concentrations (~0.5 mM) consistently promote higher RT fidelity compared to standard conditions (5-10 mM Mg²⁺), explaining longstanding discrepancies between in vitro and cellular mutation rates. These findings necessitate reconsideration of established RT characterization protocols and antiretroviral drug screening methodologies. Researchers should implement physiological Mg²⁺ conditions when seeking biologically relevant insights into RT function, particularly for fidelity assessment and therapeutic development.
Strategic Optimization of Magnesium Chloride for Diverse PCR Applications
In the polymerase chain reaction (PCR), every component must be precisely balanced to achieve specific and efficient amplification of DNA. Among these, magnesium chloride (MgCl₂) is not merely a buffer component but a critical cofactor that directly governs the activity of DNA polymerase and the fidelity of the entire reaction [1] [2]. While other reagents provide the building blocks or blueprint for amplification, Mg²⁺ ions act as the fundamental catalyst. Establishing an optimal Mg²⁺ concentration is therefore a prerequisite for successful PCR, especially in demanding applications like diagnostics and drug development. This guide establishes the standard Mg²⁺ concentration range of 1.5–2.0 mM, explores the biochemical rationale behind it, and provides a framework for comparing reaction performance against this baseline, equipping researchers with the data needed to evaluate PCR fidelity.
The Fundamental Role of Mg²⁺ in PCR
The magnesium ion (Mg²⁺) is a divalent cation that performs two non-negotiable functions in the PCR process, both crucial for the formation of new DNA strands.
Polymerase Cofactor: At the heart of the replication machinery, Mg²⁺ is an essential cofactor for Taq DNA polymerase and other PCR enzymes [1] [2]. The ion binds directly to the enzyme's active site, where it facilitates the formation of a phosphodiester bond between the 3′-OH group of the primer and the phosphate group of the incoming deoxynucleoside triphosphate (dNTP) [2]. Without Mg²⁺, the polymerase exhibits dramatically reduced, or even absent, catalytic activity [1].
Nucleic Acid Stabilizer: Beyond its role with the enzyme, Mg²⁺ is vital for the stability of the primer-template complex. The DNA backbone is highly negatively charged due to its phosphate groups, creating electrostatic repulsion that can prevent a primer from binding effectively to its complementary template strand. Mg²⁺ ions neutralize this repulsion by binding to the phosphate backbone, thereby facilitating proper annealing and increasing the melting temperature (Tm) of the duplex [1]. This dual role makes Mg²⁺ concentration a primary lever for controlling both the efficiency and the specificity of the PCR.
Quantitative Analysis: Establishing the Standard 1.5–2.0 mM Range
The consensus within the molecular biology community, supported by extensive empirical data, points to a standard Mg²⁺ concentration range of 1.5 to 2.0 mM for routine PCR applications using Taq DNA Polymerase [28]. This range is not arbitrary but represents a balance that supports robust amplification while minimizing errors.
Supporting Data from Guidelines and Meta-Analyses
A meta-analysis of 61 peer-reviewed studies confirmed that MgCl₂ concentration has a direct, measurable impact on PCR thermodynamics, with an optimal range identified between 1.5 and 3.0 mM [5]. The analysis further quantified that every 0.5 mM increase in MgCl₂ within this range is associated with a 1.2 °C increase in DNA melting temperature [5], directly illustrating how Mg²⁺ stabilizes nucleic acid duplexes. Independent guidelines from New England Biolabs (NEB), a leading enzyme manufacturer, explicitly recommend 1.5-2.0 mM Mg²⁺ as optimal for Taq DNA Polymerase [28]. This range is designed to provide sufficient cofactor for the polymerase while accounting for the chelation of Mg²⁺ by other reaction components like dNTPs and the DNA template itself [28].
The table below summarizes the effects of Mg²⁺ concentration on PCR outcomes, providing a clear baseline for comparison.
Table 1: The Impact of Magnesium Chloride Concentration on PCR Performance
| Mg²⁺ Concentration | Expected Outcome on PCR | Observed Gel Electrophoresis Result | Primary Rationale |
|---|---|---|---|
| Too Low (< 1.5 mM) | Weak or no amplification [28] [1]. | Faint product band or no band; possible smearing [7]. | Insufficient polymerase cofactor activity; poor primer annealing stability [1]. |
| Optimal (1.5 - 2.0 mM) | Specific and efficient amplification of desired product [28]. | A single, sharp band of the expected size. | Balanced conditions for high polymerase activity and specific primer-template binding. |
| Too High (> 2.0 mM) | Non-specific amplification; possible primer-dimer formation [28] [1]. | Multiple bands or a smear of non-specific products [7]. | Reduced reaction stringency promotes mis-priming at non-target sites [1]. |
The Cofactor-dNTP Equilibrium
A key rationale for the 1.5–2.0 mM range lies in the essential interaction between Mg²⁺ and dNTPs. The Mg²⁺ ion used catalytically by the polymerase is bound to a dNTP molecule, forming a Mg-dNTP complex that is the true substrate for the enzyme [1]. Because dNTPs chelate Mg²⁺, the free concentration of Mg²⁺ available to the polymerase is the total concentration minus that bound to dNTPs. A standard dNTP mix (e.g., 200 µM of each dNTP) chelates a significant amount of Mg²⁺. The 1.5–2.0 mM range is calculated to provide an excess of free Mg²⁺ after this chelation, ensuring the enzyme functions at peak efficiency without promoting the error-prone binding associated with high concentrations.
Experimental Protocols for Determining Optimal Mg²⁺
To move from the theoretical baseline to a validated, optimized protocol for a specific assay, empirical testing is required. The following methodology provides a robust framework for this optimization.
Standardized Mg²⁺ Titration Protocol
This protocol is adapted from common laboratory practices and peer-reviewed optimization studies [29] [28] [30].
- Reaction Setup: Prepare a master mix containing all standard PCR components: 1X PCR buffer (without Mg²⁺), 0.2 µM of each primer, 200 µM of each dNTP, 0.5–2.5 units of Taq DNA Polymerase, template DNA (e.g., 5–50 ng genomic DNA), and nuclease-free water [30].
- Mg²⁺ Titration: Aliquot the master mix into separate PCR tubes. Supplement each tube with MgCl₂ from a stock solution (e.g., 25 mM) to create a final concentration gradient. A standard titration series is 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, and 4.0 mM [28].
- Thermal Cycling: Run the reactions using standard cycling conditions appropriate for the primer pair and template. An initial denaturation at 95°C for 2 minutes, followed by 25–35 cycles of denaturation (95°C for 15–30 s), annealing (Ta for 15–30 s), and extension (68°C for 1 min/kb), with a final extension at 68°C for 5–10 minutes is typical [28] [30].
- Analysis: Resolve the PCR products by agarose gel electrophoresis. The optimal Mg²⁺ concentration is identified as the lowest concentration that produces a single, intense band of the correct amplicon size without non-specific bands or smearing [29].
Case Study: Optimizing a GC-Rich EGFR Promoter Amplification
A study aiming to amplify the GC-rich promoter region of the EGFR gene (75.45% GC content) from formalin-fixed paraffin-embedded (FFPE) tissue provides a clear example of moving beyond the standard baseline [29]. The researchers systematically tested MgCl₂ concentrations from 0.5 to 2.5 mM. While the standard 1.5–2.0 mM range was a good starting point, they found that 1.5 mM MgCl₂ was the specific optimum for their challenging template when combined with other optimizers like 5% DMSO [29]. This highlights that while the standard range is an essential baseline, precise optimization is key for difficult templates, and the optimal concentration can lie at the lower end of the standard window.
Table 2: Research Reagent Solutions for PCR Optimization
| Item | Function in Mg²⁺ Optimization | Example in Protocol |
|---|---|---|
| Taq DNA Polymerase | The core enzyme whose activity is directly dependent on Mg²⁺ as a cofactor. | 0.5–2.5 units per 50 µL reaction [30]. |
| dNTP Mix | Building blocks for DNA synthesis; chelate Mg²⁺ and determine free [Mg²⁺] available. | 200 µM of each dNTP is standard; concentration affects required [Mg²⁺] [28]. |
| MgCl₂ Stock Solution | The source of Mg²⁺ ions for the reaction; allows for precise concentration titration. | Typically a 25 mM stock used to create a gradient from 0.5 to 4.0 mM [28]. |
| PCR Buffer (without Mg²⁺) | Provides the chemical environment (pH, salts); using a Mg-free buffer is essential for a clean titration. | 1X concentration; often supplied as a 10X stock by the manufacturer [30]. |
| Additives (e.g., DMSO) | Can help amplify difficult templates (GC-rich, high secondary structure), interacting with Mg²⁺ optimization. | 5% DMSO was critical for successful EGFR promoter amplification [29]. |
Comparative Performance: Standard vs. Suboptimal Concentrations
Evaluating PCR fidelity requires a direct comparison of outputs across the Mg²⁺ gradient. The performance differential between the standard optimal range and suboptimal concentrations is stark and directly measurable.
Fidelity and Specificity: At the optimal 1.5–2.0 mM range, the reaction achieves high fidelity and specificity. The polymerase is fully active, and the stringency is sufficient to ensure primers anneal only to their perfect complementary sequences. This results in a single, clean band on a gel, confirming a homogeneous population of the desired amplicon [28]. In contrast, high Mg²⁺ concentrations (>2.0 mM) reduce stringency, leading to non-specific amplification. This manifests as multiple bands or a smear on an agarose gel, indicating a mixture of incorrect products and compromising downstream analyses [7] [1].
Yield and Efficiency: Mg²⁺ concentrations below 1.5 mM directly inhibit polymerase activity, leading to drastically reduced yield or complete PCR failure [28] [1]. The low ion availability results in incomplete primer extension and weak product formation, which can appear as a faint band or a smear of incomplete fragments [7]. The optimal range provides the highest consistent yield of the correct product, which is critical for applications like cloning, sequencing, and clinical diagnostics where output quantity and purity are paramount.
The establishment of a 1.5–2.0 mM Mg²⁺ baseline is a cornerstone of reliable PCR protocol design. This range is underpinned by a clear biochemical rationale that balances the cofactor requirements of the DNA polymerase with the need for stable, specific primer-template binding. For researchers evaluating PCR fidelity, this baseline serves as the essential control against which alternative conditions must be compared. While the standard range is sufficient for many applications, this guide demonstrates that a systematic titration is a non-negotiable step for assay development, particularly with challenging templates like GC-rich sequences or clinically derived FFPE samples. By applying the standardized protocols and comparative framework outlined here, scientists in drug development and basic research can make data-driven decisions to ensure their PCR results are both specific and reproducible, forming a solid foundation for their scientific conclusions.
In polymerase chain reaction (PCR) optimization, magnesium chloride (MgCl₂) concentration serves as a pivotal cofactor that fundamentally influences enzyme kinetics, primer-template binding efficiency, and ultimately, the fidelity and specificity of amplification. As a necessary cofactor for thermostable DNA polymerases, Mg²⁺ ions facilitate the formation of catalytically active enzyme complexes and stabilize the interaction between primers and template DNA. The systematic titration of MgCl₂ from 0.5 mM to 5.0 mM provides a methodological framework for identifying optimal reaction conditions that maximize amplification efficiency while minimizing non-specific products. This investigation is particularly relevant for researchers aiming to establish robust PCR protocols for diagnostic applications, gene expression analysis, and amplification of challenging templates, including GC-rich sequences.
Recent meta-analyses of PCR optimization have quantitatively demonstrated that template characteristics significantly influence MgCl₂ requirements, with genomic DNA templates typically requiring higher concentrations than simpler plasmid DNA templates [5]. The interplay between Mg²⁺ concentration and DNA melting temperature follows a predictable logarithmic relationship, enabling researchers to strategically modulate reaction stringency through controlled alterations of magnesium concentration. Within the context of evaluating PCR fidelity across different magnesium concentrations, this experimental approach provides the empirical foundation necessary for establishing reproducible and reliable amplification conditions across diverse laboratory settings and template types.
Theoretical Framework: Mg²⁺ in Reaction Thermodynamics and Kinetics
Biochemical Mechanisms of Magnesium in PCR
Magnesium ions play multifunctional roles in PCR that collectively impact reaction thermodynamics and enzymatic fidelity. Primarily, Mg²⁺ serves as an essential cofactor for DNA polymerase activity by facilitating the binding of deoxynucleotide triphosphates (dNTPs) to the enzyme's active site and catalyzing the phosphodiester bond formation between the 3'-hydroxyl group of the primer and the α-phosphate group of the incoming nucleotide [31]. This fundamental biochemical role makes Mg²⁺ concentration a direct determinant of polymerase processivity and catalytic efficiency. Additionally, Mg²⁺ stabilizes the double-stranded structure of DNA by neutralizing the negative charges on the phosphate backbone, thereby reducing electrostatic repulsion between primer and template strands and influencing the melting temperature (Tm) of the duplex.
Quantitative analyses have established that MgCl₂ concentration exhibits a logarithmic relationship with DNA melting temperature, with every 0.5 mM increase in MgCl₂ within the 1.5-3.0 mM range associated with an approximate 1.2°C increase in melting temperature [5]. This thermodynamic influence directly impacts primer annealing efficiency and specificity, particularly for templates with complex secondary structures or high GC content. The stabilization effect also extends to the final PCR product, where insufficient Mg²⁺ can lead to reduced yield due to incomplete primer extension, while excess Mg²⁺ may promote non-specific amplification by stabilizing mismatched primer-template interactions.
Concentration-Dependent Effects on PCR Performance Parameters
The optimization of MgCl₂ concentration represents a critical balance between reaction specificity and efficiency, with distinct concentration-dependent effects observed across the 0.5-5.0 mM range. At suboptimal concentrations (<1.0 mM), DNA polymerase activity becomes significantly impaired due to insufficient cofactor availability, resulting in reduced processivity and low product yield. Conversely, supraoptimal concentrations (>4.0 mM) diminish reaction stringency by stabilizing non-specific primer-template interactions, potentially leading to spurious amplification products and primer-dimer formation [18].
Table 1: MgCl₂ Concentration Effects on PCR Performance Parameters
| MgCl₂ Concentration (mM) | Polymerase Activity | Reaction Specificity | Typical Application |
|---|---|---|---|
| 0.5 - 1.0 | Significantly reduced | High | High-specificity targets; simple templates |
| 1.5 - 2.0 | Optimal | High | Standard PCR; plasmid DNA |
| 2.5 - 3.0 | High | Moderate | Genomic DNA; complex templates |
| 3.5 - 4.0 | High | Reduced | GC-rich regions; problematic amplicons |
| 4.5 - 5.0 | High | Significantly reduced | Special applications only |
Template characteristics significantly influence the optimal MgCl₂ concentration, with genomic DNA templates typically requiring approximately 0.5-1.0 mM higher concentrations than plasmid templates due to their increased complexity and potential for secondary structure formation [5]. This template-specific response underscores the necessity of empirical optimization through systematic titration experiments, particularly when working with novel target sequences or suboptimal primer designs.
Experimental Design: MgCl₂ Titration Methodology
Reagent Preparation and Standardization
The implementation of a methodical MgCl₂ titration experiment requires careful preparation and standardization of all reaction components to ensure reproducible results. A master mix approach is recommended to minimize tube-to-tube variation, with the MgCl₂ component added separately to create the concentration gradient. This methodology ensures consistent concentrations of all other reaction components while systematically varying only the magnesium parameter.
Table 2: Research Reagent Solutions for MgCl₂ Titration Experiments
| Reagent | Function in PCR | Standard Concentration | Optimization Considerations |
|---|---|---|---|
| DNA Polymerase | Enzymatic DNA synthesis | 0.5-2.5 U/50 μL reaction | Choice affects fidelity & efficiency; inhibitor-tolerant variants available [32] |
| dNTPs | DNA synthesis substrates | 200 μM each | Balanced concentration critical; affects free Mg²⁺ availability |
| Primers | Target sequence recognition | 0.2-1.0 μM each | Higher concentrations may increase non-specific binding [18] |
| Buffer Components | Maintain pH and ionic strength | 10-50 mM Tris-HCl, 50 mM KCl | Provides reaction stability; compatible with enzyme |
| Template DNA | Amplification target | 1 ng-1 μg | Quality affects optimization; inhibitor-free preparation ideal |
| MgCl₂ | Enzyme cofactor; stabilizer | Variable: 0.5-5.0 mM | Parameter under investigation |
For the titration series, prepare a stock MgCl₂ solution at a standardized concentration (typically 25-50 mM) to enable precise pipetting while minimizing volumetric errors. The remaining master mix components should be prepared at 1.1-1.25× the final desired concentration to account for the dilution factor introduced by the MgCl₂ stock solution and template DNA. The use of nuclease-free water and high-purity reagents is essential to prevent contamination from alternative sources of divalent cations that might confound results.
Titration Protocol and Thermal Cycling Parameters
The experimental workflow for MgCl₂ titration follows a systematic approach to generate comparable amplification results across the concentration gradient. The following protocol is adapted from established PCR optimization methodologies with specific modifications for magnesium concentration screening [18] [5].
Procedure:
- Master Mix Preparation: Combine all reaction components except MgCl₂ and template DNA in a 1.5 mL microcentrifuge tube according to the proportions outlined in Table 3. Mix thoroughly by gentle vortexing followed by brief centrifugation.
- Aliquoting: Dispense equal volumes of the master mix into ten labeled PCR tubes or a 96-well PCR plate.
- MgCl₂ Addition: Add the appropriate volume of MgCl₂ stock solution to each tube to create the concentration gradient from 0.5 mM to 5.0 mM in 0.5 mM increments.
- Template Addition: Add an equal quantity of template DNA to each reaction tube, using a standardized amount appropriate for the specific application (typically 1-100 ng for genomic DNA).
- Thermal Cycling: Program the thermal cycler with the parameters outlined below and initiate amplification.
Table 3: Sample Reaction Setup for MgCl₂ Titration (50 μL total volume)
| Component | Stock Concentration | Final Concentration | Volume per Reaction (μL) |
|---|---|---|---|
| 10× PCR Buffer | 10× | 1× | 5.0 |
| dNTP Mix | 10 mM each | 200 μM each | 1.0 |
| Forward Primer | 10 μM | 0.5 μM | 2.5 |
| Reverse Primer | 10 μM | 0.5 μM | 2.5 |
| Template DNA | Variable | 1-100 ng | Variable |
| MgCl₂ | 25 mM | 0.5-5.0 mM | 1.0-10.0 |
| DNA Polymerase | 5 U/μL | 1.25 U | 0.25 |
| Nuclease-Free Water | - | - | To 50 μL |
Thermal Cycler Program:
- Initial Denaturation: 95°C for 2-5 minutes
- Amplification Cycles (30-35 cycles):
- Denaturation: 95°C for 30 seconds
- Annealing: Primer-specific temperature (typically 55-65°C) for 30 seconds
- Extension: 72°C for 1 minute per kilobase of expected product
- Final Extension: 72°C for 5-10 minutes
- Hold: 4-10°C
Data Analysis and Interpretation
Quantitative and Qualitative Assessment Methods
Following amplification, systematic analysis of PCR products across the MgCl₂ concentration gradient enables comprehensive assessment of reaction performance. Agarose gel electrophoresis (1-2% agarose in TAE or TBE buffer) provides initial qualitative information regarding product specificity, yield, and the presence of non-specific amplification artifacts. For quantitative comparisons, real-time PCR systems with intercalating dyes (e.g., SYBR Green I) offer precise measurements of amplification efficiency and reaction kinetics through quantification cycle (Cq) values [12].
The optimal MgCl₂ concentration is identified as the lowest concentration that generates robust, specific amplification of the target sequence without secondary products. This concentration typically corresponds to reactions displaying single, intense bands of expected size on agarose gels, minimal primer-dimer formation, and the lowest Cq values in quantitative assays. Documentation should include gel images with appropriate molecular weight standards and quantitative measurements of band intensity or fluorescence kinetics to support objective comparison.
Troubleshooting Common Optimization Challenges
Several common amplification issues may arise during MgCl₂ optimization that require specific interpretive approaches:
- No Amplification Across All Concentrations: This may indicate primer design issues, template quality problems, or insufficient polymerase activity. Verify template quality and primer specificity before repeating titration.
- Smearing or Multiple Bands at Higher Concentrations: Excessive Mg²⁺ (>3.0-4.0 mM) typically reduces reaction stringency, leading to non-specific priming. The optimal concentration will be lower in this range.
- Reduced Yield at Intermediate Concentrations: This unexpected pattern may indicate Mg²⁺-dependent secondary structure formation or enzyme inhibition. Consider incorporating additives such as DMSO, betaine, or commercial GC enhancers that modify DNA melting behavior [31].
- Inconsistent Replicates: Technical variation often stems from pipetting errors with small volumes of MgCl₂ stock solutions. Prepare a dilution series of the MgCl₂ stock to improve pipetting accuracy for low-concentration reactions.
For templates with particularly challenging characteristics, such as high GC content (>60%), consider extending the titration range or incorporating specialized polymerase systems formulated for difficult templates. Research indicates that GC-rich templates often benefit from marginally higher MgCl₂ concentrations (3.0-4.5 mM) combined with additives that reduce secondary structure formation [31].
Comparative Performance Across Polymerase Systems
Enzyme-Specific Magnesium Requirements
The optimal MgCl₂ concentration exhibits significant variation across different DNA polymerase systems due to their distinct biochemical properties and buffer formulations. While standard Taq DNA polymerase typically functions optimally at 1.5-2.0 mM MgCl₂, engineered variants and high-fidelity enzymes may demonstrate altered magnesium dependencies. For instance, inhibitor-tolerant polymerase formulations maintain activity across a broader magnesium range, enhancing protocol robustness when working with complex sample matrices [32].
Table 4: Comparative MgCl₂ Optimization Across DNA Polymerase Systems
| DNA Polymerase Type | Typical Optimal [MgCl₂] Range | Fidelity Relative to Taq | Considerations for MgCl₂ Titration |
|---|---|---|---|
| Standard Taq | 1.5-2.5 mM | 1× | Baseline for comparison; sensitive to concentration changes |
| High-Fidelity (e.g., Q5) | 1.0-2.5 mM | 280× | May require narrower optimization range [31] |
| Inhibitor-Tolerant (e.g., StellarTaq) | 2.0-3.0 mM | Similar to Taq | Tolerant to broader concentration range [32] |
| Blended Systems | 1.5-3.0 mM | Variable | May require specific buffer formulations |
| Fast Cycling Formats | 2.0-3.0 mM | Similar to Taq | Often optimized for speed over fidelity |
When transitioning between polymerase systems, re-optimization of MgCl₂ concentration is strongly recommended, as buffer composition and enzyme kinetics significantly influence magnesium requirements. Commercial polymerase formulations often include proprietary buffer systems pre-optimized for specific magnesium concentrations; deviation from these recommended conditions may necessitate comprehensive re-optimization to maintain amplification efficiency and specificity.
Application-Specific Optimization Strategies
The methodological approach to MgCl₂ titration should be tailored to specific PCR applications and template characteristics:
- Quantitative PCR (qPCR): Optimal MgCl₂ concentration critically influences amplification efficiency, which directly impacts quantification accuracy. The MIQE guidelines emphasize the importance of reporting optimization procedures and reaction conditions to ensure reproducible results [33] [34].
- High-Resolution Melting (HRM) Analysis: Magnesium concentration affects duplex stability and melting temperature, requiring precise optimization for accurate genotype discrimination [35] [36].
- Multiplex PCR: Simultaneous amplification of multiple targets typically requires compromised MgCl₂ concentrations that support efficient amplification of all targets, often slightly higher than standard single-plex reactions.
- RT-PCR and One-Step RT-qPCR: Magnesium requirements may differ significantly due to the presence of reverse transcriptase and its buffer components, with evidence that some Taq polymerase variants exhibit tolerance to these conditions [12] [32].
For all applications, proper documentation of optimization procedures following established reporting guidelines such as MIQE ensures experimental transparency and methodological reproducibility [33]. This practice is particularly important when evaluating PCR fidelity across different magnesium concentrations as part of broader thesis research.
Systematic MgCl₂ titration from 0.5 mM to 5.0 mM provides a fundamental methodology for establishing robust, specific, and efficient PCR amplification conditions across diverse experimental contexts. The empirical determination of optimal magnesium concentration accounts for template-specific characteristics, primer design parameters, and enzyme-specific requirements that collectively influence reaction performance. Through the implementation of this standardized titration approach, researchers can methodically identify conditions that maximize target specificity while minimizing artifacts, thereby enhancing the reliability and reproducibility of molecular analyses. The integration of these optimization data with established reporting frameworks advances methodological rigor in PCR-based research and diagnostic applications.
The optimization of magnesium ion (Mg²⁺) concentration represents a fundamental parameter in polymerase chain reaction (PCR) that directly influences amplification success, particularly when challenging templates are involved. Within the context of PCR fidelity research, Mg²⁺ serves not merely as a reaction component but as a central coordinator of multiple biochemical interactions that determine reaction outcome. As a required cofactor for thermostable DNA polymerases, Mg²⁺ enables enzyme activity through catalytic facilitation of phosphodiester bond formation and stabilization of the primer-template complex [2] [37]. However, its concentration must be precisely calibrated according to template characteristics, as inappropriate Mg²⁺ levels can dramatically reduce enzyme fidelity and increase nonspecific amplification [37].
This guide systematically evaluates template-specific Mg²⁺ optimization strategies across three challenging amplification scenarios: GC-rich sequences, long amplicons, and complex genomic DNA. By comparing experimental data across multiple studies and polymerase systems, we provide evidence-based protocols for researchers seeking to maximize amplification efficiency while maintaining fidelity in molecular diagnostics, genetic research, and drug development applications.
Biochemical Mechanisms: How Mg²⁺ Influences PCR Fidelity
Molecular Interactions at the Polymerase Active Site
The coordination of Mg²⁺ at the DNA polymerase active site follows a well-established mechanistic model wherein two magnesium ions participate directly in the catalytic transfer of the phosphoryl group during nucleotide incorporation [2]. The first metal ion (Metal A) activates the 3'-hydroxyl group of the primer terminus for nucleophilic attack, while the second (Metal B) facilitates departure of the pyrophosphate group from the incoming dNTP. This precise geometric arrangement creates a transition state that significantly accelerates the rate of phosphodiester bond formation—without Mg²⁺, DNA polymerases exhibit minimal catalytic activity [2] [38].
Beyond its role in catalysis, Mg²⁺ serves essential structural functions throughout the PCR process. The cation stabilizes the double-stranded DNA structure by neutralizing the negative charges on the phosphate backbone, thereby reducing electrostatic repulsion between primer and template strands [2] [39]. This charge screening effect lowers the thermodynamic barrier to hybridization, facilitating primer binding particularly in sequences with high secondary structure potential. Research demonstrates that Mg²⁺ concentration directly influences DNA duplex stability, with a 1 M increase in MgCl₂ concentration raising the melting temperature (Tm) by approximately 5.5°C [38].
Fidelity Implications of Mg²⁺ Concentration
The relationship between Mg²⁺ concentration and polymerase fidelity follows a U-shaped curve, wherein both deficiency and excess reduce accuracy. At suboptimal concentrations, inadequate Mg²�+ availability slows catalytic rate and can promote polymerase stalling, particularly through structured regions. Conversely, excess Mg²⁺ reduces enzyme fidelity by relaxing base-pairing stringency and increasing misincorporation rates [37]. Recent investigations into reverse transcriptase fidelity have revealed that physiological Mg²⁺ concentrations (approximately 0.5 mM) promote higher fidelity compared to the elevated concentrations (5-10 mM) traditionally used in vitro optimization protocols [25].
Table 1: Comparative Fidelity of DNA Polymerases Under Different Mg²⁺ Concentrations
| Polymerase Type | Low Mg²⁺ (0.5-1 mM) | Standard Mg²⁺ (1.5-2 mM) | High Mg²⁺ (3-4 mM) | Primary Application |
|---|---|---|---|---|
| Standard Taq | Reduced activity | Balanced fidelity/yield | Increased errors | Routine amplification |
| Proofreading | High fidelity | Moderate fidelity | Reduced fidelity | Cloning, sequencing |
| GC-optimized | Often insufficient | Target-specific optimization | Increased nonspecific | GC-rich targets |
Template-Specific Mg²⁺ Optimization Strategies
GC-Rich Templates
GC-rich templates (≥60% GC content) present unique challenges for PCR amplification due to their propensity for forming stable secondary structures and their elevated melting temperatures. The strong hydrogen bonding between G-C base pairs (three bonds versus two in A-T pairs) creates regions that resist denaturation and promote primer misannealing [39]. Research on the epidermal growth factor receptor (EGFR) promoter sequence, which features extremely high GC content up to 88%, demonstrated that successful amplification required precise MgCl₂ optimization between 1.5-2.0 mM [29].
In this study, which aimed to amplify a 197-bp fragment containing two SNPs (-216G>T and -191C>A) from formalin-fixed paraffin-embedded lung tumor tissue, investigators systematically optimized reaction conditions. They found that MgCl₂ concentrations below 1.5 mM resulted in weak or no amplification, while concentrations exceeding 2.0 mM promoted nonspecific products. The optimized protocol incorporated 5% dimethyl sulfoxide (DMSO) as a necessary additive to destabilize secondary structures, while the annealing temperature required elevation to 63°C—7°C higher than calculated based on primer Tm alone [29].
Table 2: Comprehensive Optimization Protocol for GC-Rich Templates
| Parameter | Suboptimal Condition | Optimized Condition | Rationale |
|---|---|---|---|
| MgCl₂ | 0.5-1.0 mM or >2.5 mM | 1.5-2.0 mM | Balances polymerase activity with primer specificity |
| Additives | None | 5% DMSO or betaine | Disrupts secondary structures, lowers actual Tm |
| Annealing Temperature | Calculated Tm | Tm + 5-10°C | Counteracts reduced stringency from additives |
| DNA Concentration | <1.86 μg/mL | ≥2 μg/mL | Provides sufficient template despite reduced accessibility |
| Polymerase Choice | Standard Taq | GC-optimized enzymes | Better progression through structural barriers |
For particularly stubborn GC-rich targets, empirical testing through Mg²⁺ gradient PCR (0.5 mM increments between 1.0-4.0 mM) is recommended [39]. Commercial polymerase systems specifically formulated for GC-rich amplification, such as Q5 High-Fidelity DNA Polymerase with GC Enhancer, provide pre-optimized buffer conditions that mitigate the need for extensive optimization [39].
Long Amplicons
Amplification of long DNA fragments (>5 kb) introduces distinct challenges that demand specialized Mg²⁺ optimization strategies. The extended elongation times required for long amplicons increase the probability of polymerase dissociation or enzymatic errors. Additionally, the statistical likelihood of encountering problematic secondary structures or regions of extreme GC content increases with amplicon length.
For long amplicon amplification, Mg²⁺ concentrations typically require elevation to stabilize the polymerase-template complex throughout extended elongation phases. While standard PCR may function optimally at 1.5-2.0 mM MgCl₂, long-range protocols often benefit from concentrations between 2.0-2.5 mM [2]. However, this elevation must be carefully balanced against the fidelity reduction associated with excess Mg²⁺, particularly when using non-proofreading polymerases.
The experimental protocol for long amplicon amplification should incorporate a stepped Mg²⁺ optimization approach, beginning with the manufacturer's recommended concentration for the selected long-range polymerase and testing 0.25 mM increments above and below this baseline. Extension times must be calibrated according to polymerase processivity—approximately 1-2 minutes per kilobase for most modern engineered enzymes. A touchdown PCR approach, with initial high annealing temperatures that gradually decrease over subsequent cycles, can improve specificity for long targets while working in concert with optimized Mg²⁺ concentrations [40].
Complex Genomic DNA
Complex genomic DNA templates, particularly those derived from suboptimal sources such as formalin-fixed paraffin-embedded (FFPE) tissues or forensic samples, present compound challenges including cross-linking, fragmentation, and co-purification of inhibitors. The Mg²⁺ requirements for these templates often deviate significantly from standard recommendations due to both template compromise and the presence of confounding factors.
Research on FFPE-derived DNA demonstrates that formalin fixation causes formation of nucleoprotein complexes and methylene bridging between nucleic acids and proteins, reducing template accessibility [29]. In these cases, slightly elevated Mg²⁺ concentrations (2.0-2.5 mM) may help stabilize the polymerase on compromised templates. However, the co-purification of divalent cation chelators from fixation processes may effectively reduce free Mg²⁺ concentration, necessitating empirical determination of optimal conditions.
For complex genomic DNA, a sequential optimization approach is recommended:
- Quantify DNA integrity through electrophoretic analysis and spectrophotometric ratios (A260/A280 ~1.8, A260/A230 >1.8) [40]
- Perform inhibitor assessment via spiked control reactions
- Titrate MgCl₂ across a broad range (1.0-4.0 mM in 0.5 mM increments)
- Evaluate additive enhancement with DMSO, betaine, or commercial GC enhancers
This protocol successfully enabled amplification of the challenging EGFR promoter region from FFPE tissue, demonstrating the critical importance of template-specific optimization when working with complex genomic DNA [29].
Mathematical Modeling for Mg²⁺ Prediction
Recent advances in predictive modeling have enabled more precise estimation of optimal Mg²⁺ concentrations based on reaction parameters. A 2025 study developed a multivariate Taylor series expansion model that incorporates thermodynamic principles to predict MgCl₂ requirements with high accuracy (R² = 0.9942) [38].
The foundational relationship expresses MgCl₂ concentration as a function of multiple variables: (MgCl₂) = f (Tm, GC%, L, (dNTP), (Primers), (Polymerase), pH, T)
This was expanded to a sophisticated third-order model: (MgCl₂) = β₀ + Σᵢβᵢxᵢ + Σᵢ Σⱼ βᵢⱼxᵢxⱼ + Σᵢ Σⱼ Σₖ βᵢⱼₖxᵢxⱼxₖ + βL ln(L) + βH(ΔH/RT) + β_S(ΔS/R) + ε
The resulting predictive equation identified key factors influencing Mg²⁺ optimization: (MgCl₂) ≈ 1.5625 + (-0.0073 × Tm) + (-0.0629 × GC) + (0.0273 × L) + (0.0013 × dNTP) + (-0.0120 × Primers) + (0.0007 × Polymerase) + (0.0012 × log(L)) + (0.0016 × TmGC) + (0.0639 × dNTPPrimers) + (0.0056 × pH_Polymerase)
Variable importance analysis revealed that the interaction between dNTP and primers accounted for 28.5% of predictive power, followed by GC content (22.1%) and amplicon length (15.7%) [38]. This model provides a theoretical framework for initial Mg²⁺ estimation before empirical optimization, potentially reducing optimization time for novel targets.
Comparative Experimental Data
Systematic Mg²⁺ Titration Across Template Types
Experimental data from multiple studies enables direct comparison of Mg²⁺ optimization requirements across template categories. A systematic investigation of the EGFR promoter region (75.45% GC content) demonstrated absolute requirement for 1.5-2.0 mM MgCl₂, with no amplification observed outside this range under otherwise optimized conditions [29]. This narrow optimal window contrasts with the broader tolerance (1.0-4.0 mM) observed for standard templates, highlighting the heightened Mg²⁺ sensitivity of GC-rich targets.
Table 3: Comparative Mg²⁺ Optimization Across Template Types
| Template Category | Optimal [MgCl₂] | Critical Parameters | Recommended Additives | Annealing Temperature Adjustment |
|---|---|---|---|---|
| Standard | 1.5-2.0 mM | Primer Tm, complexity | None typically | Calculated Tm -5°C |
| GC-rich | 1.5-2.0 mM | GC%, secondary structure | DMSO, betaine, 7-deaza-dGTP | Tm +5-10°C |
| Long amplicons | 2.0-2.5 mM | Length, polymerase processivity | BSA, commercial enhancers | Stepped or touchdown |
| Complex genomic | 1.5-3.0 mM | Purity, fragmentation, inhibitors | Proteinase K, BSA | Empirical determination |
Research comparing fidelity across Mg²⁺ concentrations revealed that physiological Mg²⁺ levels (∼0.5 mM) promote higher fidelity in HIV-1 reverse transcriptase compared to standard in vitro concentrations (6 mM) [25]. This fidelity effect was polymerase-specific, with Moloney murine leukemia virus (MuLV) and avian myeloblastosis virus (AMV) RTs showing equivalent fidelity across Mg²⁺ concentrations, while prototype foamy virus (PFV) RT demonstrated higher fidelity at lower Mg²⁺ [25]. These findings highlight the importance of considering both template characteristics and polymerase properties when optimizing Mg²⁺ for fidelity-critical applications.
Experimental Workflow for Mg²⁺ Optimization
The following workflow provides a systematic approach for determining optimal Mg²⁺ concentrations across template types:
Detailed Methodology
Template Characterization: Determine GC content using bioinformatic tools (e.g., EMBOSS CpGPlot), assess integrity via electrophoresis, and quantify precisely using fluorometric methods [29] [40].
Polymerase Selection: Choose polymerase systems based on template challenges:
- GC-rich targets: GC-optimized enzymes with enhancers (OneTaq GC Buffer, Q5 GC Enhancer)
- Long amplicons: High-processivity systems (LA Taq, PrimeSTAR GXL)
- High-fidelity requirements: Proofreading enzymes (Q5, Phusion)
Gradient Setup: Prepare a MgCl₂ dilution series from 0.5-4.0 mM in 0.5 mM increments while maintaining constant concentration of all other components. Include both positive and negative controls.
Thermal Cycling: Employ a temperature gradient spanning at least 10°C to identify optimal annealing conditions in conjunction with Mg²⁺ concentration.
Product Analysis: Evaluate amplification success through SYBR Safe-stained agarose gels, quantifying yield and specificity. For fidelity-critical applications, supplement with sequencing analysis of products [29].
Refinement and Validation: Narrow Mg²⁺ range based on initial results and validate optimal conditions through replicate reactions and downstream application testing (sequencing, cloning, restriction analysis).
Research Reagent Solutions
Table 4: Essential Reagents for Mg²⁺ Optimization Studies
| Reagent Category | Specific Examples | Function in Optimization | Considerations |
|---|---|---|---|
| Polymerase Systems | OneTaq DNA Polymerase with GC Buffer, Q5 High-Fidelity DNA Polymerase | Provide specialized buffer formulations for challenging templates | Select based on fidelity requirements, amplification efficiency |
| Mg²⁺ Solutions | Separate MgCl₂ supplements (typically 25-50 mM stocks) | Enable precise titration of Mg²⁺ concentration | Prepare fresh, concentration-verify periodically |
| Additives | DMSO, betaine, 7-deaza-dGTP, commercial GC enhancers | Disrupt secondary structures, modify effective Tm | Concentration-dependent effects, polymerase compatibility |
| Template Standards | Control plasmids, genomic DNA standards with known characteristics | Provide benchmarking for optimization protocols | Ensure integrity, quantitative accuracy |
| Detection Reagents | SYBR Safe DNA Gel Stain, Qubit dsDNA HS Assay Kit | Enable quantification of yield and specificity | Linear detection range, compatibility with downstream applications |
The optimization of Mg²⁺ concentration for challenging PCR templates requires a sophisticated understanding of both biochemical principles and template-specific characteristics. Evidence from multiple studies demonstrates that GC-rich templates typically require 1.5-2.0 mM MgCl₂ in combination with structure-disrupting additives; long amplicons benefit from slightly elevated concentrations (2.0-2.5 mM) to stabilize polymerase processivity; while complex genomic DNA demands empirical determination across a broad range (1.0-4.0 mM) to overcome template compromise and potential inhibitors.
The development of predictive mathematical models based on thermodynamic principles represents a significant advancement in optimization methodology, potentially reducing reliance on extensive empirical testing. However, laboratory validation remains essential, particularly for applications requiring high fidelity. By integrating theoretical prediction with systematic experimental workflows, researchers can efficiently determine optimal Mg²⁺ concentrations that balance yield, specificity, and fidelity across diverse template types and application requirements.
In polymerase chain reaction (PCR) optimization, the precise relationship between magnesium ions (Mg²⁺) and deoxynucleoside triphosphates (dNTPs) represents a critical biochemical equilibrium that directly determines reaction success and fidelity. Mg²⁺ serves as an essential cofactor for DNA polymerase activity, but its availability is significantly compromised through chelation by dNTPs within the reaction milieu. This interaction creates a dynamic where "free" Mg²⁺ concentration—rather than the total amount added—dictates enzymatic efficiency. Inadequate free Mg²⁺ results in minimal polymerase activity and failed amplification, while excess free Mg²⁺ diminishes fidelity by promoting non-specific primer binding and incorporation errors [6] [37]. Understanding this chelation relationship is therefore fundamental for researchers seeking to optimize PCR conditions, particularly in applications demanding high accuracy such as cloning, sequencing, and diagnostic assay development.
The physiological relevance of Mg²⁺ concentration extends beyond mere reaction efficiency. Recent investigations into reverse transcriptase fidelity reveal that physiological free Mg²⁺ levels (approximately 0.5 mM) correlate with enhanced enzymatic accuracy for several viral polymerases, including HIV-1, compared to the higher concentrations (5-10 mM) traditionally used in optimized in vitro assays [25]. This finding underscores the importance of mimicking cellular conditions for fidelity studies and suggests that the common practice of Mg²⁺ optimization must balance maximum yield with biological relevance.
Quantitative Analysis of Mg²⁺ and dNTP Interactions
Chelation Dynamics and Stability Constants
The Mg²⁺-dNTP interaction occurs through coordinate bonding between the magnesium cation and the phosphate oxygen atoms of dNTPs, forming a stable complex that renders both components temporarily unavailable for their primary enzymatic roles. The stability of these complexes is quantified by stability constants (Ks), which vary slightly among the different dNTPs but generally fall within a high-affinity range. Experimental determinations using electron spin resonance indicate that Mg²⁺ complexes with nucleotide analogs exhibit stability constants of approximately 357-497 M⁻¹ under typical PCR conditions (pH 7.0, ionic strength 0.15 M) [41]. This substantial binding affinity means that a significant portion of added Mg²⁺ is immediately complexed upon dNTP addition, effectively reducing the concentration of free Mg²⁺ available to support DNA polymerase activity.
The practical implication of this chelation is that the dNTP concentration directly determines the baseline Mg²⁺ requirement before any free Mg²⁺ becomes available for enzymatic function. A meta-analysis of PCR optimization studies has quantified this relationship, demonstrating that each 0.2 mM increment in dNTP concentration typically chelates approximately 0.3-0.45 mM Mg²⁺ [20]. Consequently, reaction mixtures with standard dNTP concentrations (200 µM of each dNTP) require a minimum of 1.2-1.8 mM Mg²⁺ just to saturate the dNTP chelation sites before any free Mg²⁺ contributes to polymerase activation.
Comparative Effects of Mg²⁺ Concentration on PCR Performance
The table below summarizes quantitative data on how varying Mg²⁺ concentrations affect key PCR parameters, based on aggregated experimental results from multiple studies:
Table 1: Effects of Mg²⁺ Concentration on PCR Performance Parameters
| Mg²⁺ Concentration | Replication Fidelity | Reaction Efficiency | Product Specificity | Recommended Application |
|---|---|---|---|---|
| Low (0.5-1.0 mM) | Higher fidelity [25] | Failed or weak amplification [6] | High specificity | High-fidelity applications when optimized |
| Optimal (1.5-2.0 mM) | Balanced fidelity and efficiency [42] | Robust amplification | Good specificity | Routine PCR, cloning |
| High (3.0-4.5 mM) | Reduced fidelity [37] | Enhanced yield | Decreased specificity, spurious products [6] | Challenging templates |
| Excessive (>4.5 mM) | Significantly impaired fidelity | Saturated yield | Multiple non-specific bands | Not recommended |
The fidelity reduction observed at elevated Mg²⁺ concentrations has mechanistic foundations in polymerase biochemistry. Structural studies of DNA polymerases indicate that Mg²�+ ions facilitate catalysis through a two-metal-ion mechanism that positions the 3'-OH nucleophile and stabilizes the leaving pyrophosphate group [3]. When Mg²⁺ concentrations exceed the optimal range, the metal ion equilibrium shifts toward relaxed substrate discrimination, potentially enabling misincorporated nucleotides to proceed through catalysis more readily.
Experimental Approaches to Mg²⁺-dNTP Optimization
Systematic Optimization Methodologies
Determining the optimal Mg²⁺ concentration for a specific PCR application requires empirical testing across a concentration gradient while maintaining constant dNTP levels. The following workflow represents a standardized approach adopted from methodological guidelines in the literature:
Table 2: Stepwise Mg²⁺ Optimization Protocol
| Step | Procedure | Parameters | Expected Outcomes |
|---|---|---|---|
| 1. Initial Setup | Prepare MgCl₂ gradient in 0.5 mM increments | 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0 mM | Baseline for optimization |
| 2. Reaction Assembly | Keep dNTP concentration constant at 200 µM each | Total dNTPs = 800 µM | Consistent chelation across reactions |
| 3. Thermal Cycling | Apply standardized cycling conditions | Annealing temperature 5°C below primer Tm | Uniform amplification pressure |
| 4. Product Analysis | Evaluate yield and specificity via electrophoresis | Agarose gel quantification | Identification of optimal concentration range |
| 5. Refinement | Fine-tune in 0.25 mM increments around best result | ±0.5 mM from best initial result | Precision optimization |
This methodological framework proved essential in optimizing PCR amplification of GC-rich templates such as the EGFR promoter region (75.45% GC content), where researchers determined that 1.5 mM MgCl₂ in combination with 5% DMSO and an elevated annealing temperature (63°C) successfully amplified this challenging target [29]. The critical finding from this study was that optimal Mg²⁺ concentration must be determined in the context of multiple reaction parameters rather than in isolation.
Magnesium-Dependent Fidelity Assessment Protocols
Evaluating the fidelity implications of Mg²⁺ concentration requires specialized assays that measure mutation frequency rather than simple amplification yield. The lacZα-based α-complementation assay represents a robust method for quantifying polymerase error rates under different Mg²⁺ conditions [25]. This approach involves:
- Template Preparation: Using a plasmid containing the lacZα gene as an amplification target
- PCR Amplification: Conducting parallel reactions at different Mg²⁺ concentrations (e.g., 0.5 mM vs. 6.0 mM)
- Cloning and Transformation: Introducing PCR products into appropriate bacterial strains
- Mutation Screening: Identifying mutants through blue-white selection based on β-galactosidase activity
- Error Rate Calculation: Determining mutations per base pair per duplication through statistical analysis
Application of this methodology to reverse transcriptase fidelity studies demonstrated that HIV-1 RT shows approximately 4-fold higher fidelity in 0.5 mM Mg²⁺ compared to 6 mM Mg²⁺ [25]. Next Generation Sequencing (NGS) approaches further corroborated these findings, providing comprehensive mutation profiles across the entire lacZα template and offering unprecedented resolution in fidelity assessment.
Comparative Performance of PCR Enzymes Under Varied Mg²⁺ Conditions
Fidelity Profiles of Commercial Polymerases
Different DNA polymerases exhibit distinct responses to Mg²⁺ concentrations, reflecting their structural variations and catalytic mechanisms. A comprehensive comparison of six commonly used PCR enzymes under standardized conditions revealed significant fidelity differences:
Table 3: Error Rates of DNA Polymerases Under Optimal Mg²⁺ Conditions
| DNA Polymerase | Error Rate (mutations/bp/duplication) | Fidelity Relative to Taq | Optimal [Mg²⁺] Range |
|---|---|---|---|
| Taq | 3.0-5.6 × 10⁻⁵ [15] | 1x | 1.5-2.0 mM [42] |
| AccuPrime-Taq HF | ~1.0 × 10⁻⁵ [15] | ~5x better | Vendor-specific |
| KOD Hot Start | Comparable to Pfu [15] | ~50x better | Vendor-specific |
| Pfu | 1-2 × 10⁻⁶ [15] | 6-10x better | Vendor-specific |
| Phusion Hot Start | 4.0 × 10⁻⁷ (HF buffer) [15] | >50x better | 1-2 mM [37] |
| Pwo | Comparable to Pfu [15] | ~10x better | Vendor-specific |
The superior fidelity of proofreading enzymes like Pfu and Phusion persists across Mg²⁺ concentrations, but absolute error rates increase for all enzymes at elevated Mg²⁺ levels. This underscores the importance of maintaining Mg²⁺ concentrations at the lower end of the optimal range for applications requiring maximum accuracy.
Structural Basis for Metal Ion Effects on Polymerase Function
The molecular mechanisms underlying Mg²⁺ dependence in DNA polymerases have been elucidated through structural and kinetic studies. High-resolution crystal structures of DNA polymerase I (Klenow fragment) reveal that two invariant aspartate residues (Asp705 and Asp882 in Pol I(KF)) coordinate two Mg²⁺ ions at the active site [3]. Kinetic analyses of aspartate-to-alanine mutants demonstrate that:
- Asp882 serves as an anchor point for the dNTP-associated metal ion (Metal B) and is essential for the fingers-closing conformational change that creates the catalytically competent ternary complex
- Asp705 facilitates entry of the second Mg²⁺ ion (Metal A) after fingers-closing and is critical for the chemical step of nucleotide addition
- Pre-chemistry conformational steps proceed normally at very low Mg²⁺ concentrations, but higher concentrations are needed for covalent bond formation, consistent with the second metal ion entering after fingers-closing
This mechanistic understanding explains why Mg²⁺ concentration so profoundly affects both catalytic efficiency and fidelity—suboptimal concentrations impair the precise geometric arrangement of substrates and catalytic groups required for accurate nucleotide incorporation.
Research Reagent Solutions for Mg²⁺-dNTP Optimization
Successful management of the Mg²⁺-dNTP relationship requires specific laboratory reagents and materials. The following table details essential components for experimental investigation of this critical interaction:
Table 4: Essential Research Reagents for Mg²⁺-dNTP Relationship Studies
| Reagent Category | Specific Examples | Function in Optimization | Considerations |
|---|---|---|---|
| Magnesium Sources | MgCl₂, MgSO₄ | Provides essential cofactor for polymerase activity | MgCl₂ most common; concentration varies by polymerase |
| dNTP Preparations | dATP, dTTP, dCTP, dGTP | Substrates for DNA synthesis | Quality affects fidelity; concentration chelates Mg²⁺ |
| Fidelity Assay Systems | lacZα plasmid, pBS▽EcoRV567 [25] | Template for mutation frequency determination | Requires specialized cloning and expression systems |
| Polymerase Systems | Taq, Pfu, Phusion, KOD [15] | Catalyzes DNA synthesis | Fidelity and Mg²⁺ tolerance vary substantially |
| PCR Additives | DMSO, betaine, glycerol [29] | Modifies DNA melting behavior | Can affect Mg²⁺ availability and polymerase activity |
| Analysis Tools | Agarose gels, NGS platforms, restriction enzymes | Evaluates amplification success and fidelity | NGS provides most comprehensive mutation profiling |
Mg²⁺ Optimization Framework for Challenging Templates
Decision Framework for Reaction Optimization
The relationship between template characteristics and optimal Mg²⁺ concentration follows predictable patterns that can guide systematic optimization. The diagram below illustrates the decision pathway for establishing appropriate Mg²⁺ concentrations based on template properties and application requirements:
This decision framework emphasizes that GC-rich templates (≥60% GC content) generally require higher Mg²⁺ concentrations (typically 2.0-3.5 mM) to overcome template secondary structure, while applications prioritizing maximum fidelity benefit from concentrations at the lower end of the optimal range (1.5-2.0 mM) [29] [37]. The established logarithmic relationship between Mg²⁺ concentration and DNA melting temperature means that each 0.5 mM increment within the 1.5-3.0 mM range produces consistent increases in melting temperature, directly influencing reaction efficiency [20].
Advanced Considerations for Specialized Applications
For demanding applications such as long-range PCR, high-throughput cloning, or diagnostic assay development, additional factors influence the Mg²⁺-dNTP equilibrium:
- dNTP Quality and Purity: Impurities in dNTP preparations can introduce additional chelators that further reduce free Mg²⁺ availability
- Sample-Derived Chelators: Templates isolated with EDTA or citrate can introduce potent Mg²⁺ chelators that must be accounted for in reaction planning
- Polymerase-Specific Profiles: "Magnesium-tolerant" polymerases like Titanium Taq and Advantage 2 maintain activity across broader Mg²⁺ ranges (supplied with 3.5 mM MgCl₂), while high-fidelity enzymes like PrimeSTAR GXL achieve optimal performance at precisely 1 mM Mg²⁺ [37]
- Buffer Composition Variations: Commercially supplied PCR buffers often contain undefined components that may affect Mg²⁺ availability and should be held constant during optimization
Recent research on human DNA polymerase γ reveals that metal cofactors influence not only catalytic efficiency but also protein flexibility and active site stabilization, with Mg²⁺ providing greater structural stability compared to Mn²⁺ despite the latter's enhanced catalytic efficiency [43]. This trade-off between structural stability and catalytic power represents a fundamental consideration in polymerase evolution and engineering.
The critical relationship between Mg²⁺ and dNTPs represents a cornerstone of successful PCR optimization that directly connects basic biochemistry to practical experimental outcomes. The chelation equilibrium between these components dictates that they must be optimized in concert rather than independently, with the target application determining the appropriate balance point between yield and fidelity. For diagnostic applications where detection sensitivity is paramount, higher Mg²⁺ concentrations (2.5-3.5 mM) may be warranted despite modest fidelity reductions. Conversely, cloning and expression studies demand the highest possible accuracy, favoring Mg²⁺ concentrations at the lower end of the optimal range (1.5-2.0 mM) potentially supplemented with fidelity-enhancing additives like DMSO or betaine.
Future directions in Mg²⁺-dNTP relationship management will likely include polymerase engineering to reduce metal cofactor dependence, development of more sophisticated buffer systems that stabilize the free Mg²⁺ pool, and computational approaches that predict optimal conditions based on template sequence characteristics. As the field advances toward more standardized and predictable amplification systems, understanding the fundamental partnership between Mg²⁺ and dNTPs will remain essential knowledge for molecular biologists pursuing reliable genetic analysis.
Magnesium chloride (MgCl₂) serves as an essential catalytic cofactor for DNA polymerase activity, directly influencing the efficiency, specificity, and fidelity of polymerase chain reaction (PCR) [5] [44]. The concentration of Mg²⁺ impacts PCR success by modulating enzyme kinetics, primer-template binding stability, and nucleic acid melting thermodynamics [5]. Optimization is particularly crucial for advanced applications including high-fidelity cloning, diagnostic assays, and next-generation sequencing library preparation, where reaction conditions must be precisely calibrated to template characteristics [5] [45].
A recent meta-analysis of 61 peer-reviewed studies established a clear logarithmic relationship between MgCl₂ concentration and DNA melting temperature, with optimal amplification typically occurring between 1.5 and 3.0 mM [5]. Within this range, every 0.5 mM increase in MgCl₂ concentration raises the melting temperature by approximately 1.2 °C [5]. Template complexity significantly influences optimal concentration requirements, with genomic DNA templates generally requiring higher Mg²⁺ concentrations than simpler templates [5].
Experimental Methodology for Mg²⁺ Optimization
Standard Optimization Protocol
A systematic approach to Mg²⁺ optimization begins with establishing a concentration gradient across the theoretically optimal range. The following protocol adapts established methodologies from multiple experimental studies [5] [21] [44]:
- Reaction Setup: Prepare a master mix containing all PCR components except MgCl₂. Aliquot equal volumes into separate tubes.
- Concentration Gradient: Add MgCl₂ to achieve final concentrations typically ranging from 1.0 mM to 4.0 mM in 0.5 mM increments.
- Template-Specific Adjustments: For complex genomic DNA, extend the upper range to 5.0 mM; for plasmid or cDNA templates, begin as low as 0.5 mM.
- Thermal Cycling: Perform amplification using standardized cycling conditions appropriate for the specific polymerase and application.
- Product Analysis: Evaluate results using agarose gel electrophoresis for yield and specificity, and process a subset for sequencing when assessing fidelity.
Troubleshooting Common Mg²⁺-Related Issues
Common amplification problems often relate to suboptimal Mg²⁺ concentrations [7] [44]:
- Low Yield/Smearing: Typically indicates insufficient Mg²⁺, reducing polymerase activity and causing incomplete amplification [7].
- Non-Specific Bands: Often results from excessive Mg²⁺, which reduces primer-stringency and stabilizes mismatched hybrids [7] [44].
- Complete Amplification Failure: May occur at extremely low Mg²⁺ concentrations where polymerase activity is profoundly impaired [44].
Figure 1. Mg²⁺ Optimization Workflow: A systematic approach for troubleshooting PCR through magnesium concentration adjustment.
Comparative Performance Across PCR Applications
Quantitative Mg²⁺ Optimization Data
Table 1. Optimal Mg²⁺ Concentration Ranges for Various PCR Applications
| Application Type | Optimal [Mg²⁺] Range | Key Performance Metrics | Template-Specific Considerations |
|---|---|---|---|
| High-Fidelity PCR | 1.5–2.0 mM | ~50–100X Taq polymerase fidelity [45] | Lower concentrations favor accuracy; proofreading polymerases often require precise Mg²⁺ |
| Cloning & Sequencing | 1.5–2.5 mM | Minimal mutations in heterologous expression systems [45] | Blunt-end formation critical for cloning efficiency |
| Diagnostic PCR | 2.0–3.0 mM | Detection limit of 3.1 fg/µL demonstrated in fungal pathogen detection [21] | Higher concentrations may improve sensitivity for low-copy targets |
| Multiplex PCR | 2.5–3.5 mM | Simultaneous detection of 4 RNA targets with 20 copy sensitivity [23] | Balanced amplification across multiple primer sets |
| GC-Rich Templates | 2.0–3.0 mM | Success rate >90% with buffer additives [44] | Often requires complementation with DMSO or betaine |
Polymerase-Specific Magnesium Requirements
Table 2. Mg²⁺ Optimization for Different DNA Polymerase Types
| Polymerase Type | Commercial Example | Recommended [Mg²⁺] | Fidelity (vs. Taq) | Primary Applications |
|---|---|---|---|---|
| Standard Taq | Conventional Taq | 1.5–2.5 mM | 1X | Routine amplification, genotyping |
| High-Fidelity | EVO HS Super-Fidelity | 1.5–2.5 mM [45] | 50–100X [45] | Cloning, NGS, mutagenesis |
| Reverse Transcription | Engineered Taq variants | 2.0–3.5 mM [23] | Varies with mutations | Single-enzyme RT-PCR, RNA detection |
| Hot-Start | AmpliTaq Gold | 1.5–2.5 mM | 1X | Diagnostic assays, multiplexing |
Application-Specific Protocols
High-Fidelity PCR for Cloning Applications
For molecular cloning and next-generation sequencing library preparation, EVO HS Super-Fidelity DNA Polymerase exemplifies modern high-fidelity enzymes, achieving 50–100X greater accuracy than standard Taq polymerase through 3'→5' exonuclease proofreading activity [45]. The recommended reaction setup includes:
- Mg²⁺ Concentration: 1.5–2.5 mM in the proprietary reaction buffer [45]
- Thermal Cycling: Initial denaturation at 98°C for 30 seconds, followed by 30 cycles of denaturation at 98°C for 10 seconds, annealing at 55–72°C for 15–30 seconds, and extension at 72°C for 15–30 seconds/kb [45]
- Special Considerations: For GC-rich templates (>70% GC content), supplementation with DMSO (5%) or betaine (1M) is recommended to disrupt secondary structures [45] [44]
This optimized system generates blunt-end PCR products suitable for direct cloning without additional processing, making it ideal for constructing expression vectors and preparing sequencing libraries [45].
Diagnostic PCR for Pathogen Detection
In molecular diagnostics, Mg²⁺ optimization directly impacts assay sensitivity and specificity. A recent study detecting Fusarium tricinctum, a fungal pathogen of Zanthoxylum bungeanum, demonstrated that real-time quantitative PCR (qPCR) could achieve detection limits as low as 3.1 fg/µL of target DNA through meticulous reaction optimization [21]. The experimental approach included:
- Target Selection: The Fusarium-specific CYP51C gene was selected as the detection target for its ability to distinguish between closely related species [21]
- Primer Design: Specific primers were designed using Primer Premier 5.0 and validated with NCBI's Primer-BLAST tool to ensure specificity [21]
- Reaction Optimization: Systematic testing of Mg²⁺ concentrations alongside other components to establish optimal conditions for maximum sensitivity [21]
For diagnostic applications requiring extreme sensitivity, nested PCR protocols provide additional specificity through two sequential amplification rounds with inner and outer primer sets, though they require careful contamination control measures [21] [46].
Single-Enzyme Reverse Transcription PCR
Recent advancements in enzyme engineering have produced novel Taq DNA polymerase variants capable of catalyzing both reverse transcription and DNA amplification in a single tube without viral reverse transcriptases [23]. These engineered enzymes demonstrate:
- Dual Functionality: Combined reverse transcription and DNA polymerase activities enable simplified workflow [23]
- Thermostability: Retention of activity up to 95°C allows for high-temperature reverse transcription, improving specificity [23]
- Multiplexing Capability: Simultaneous detection of up to four RNA targets with a detection limit of 20 copies [23]
- Buffer Compatibility: Function with standard TaqMan probe-based detection systems without requiring Mn²⁺ additives [23]
The development of these multifunctional enzymes marks a significant advancement for molecular diagnostics, particularly for viral detection and gene expression analysis where workflow simplification and cost reduction are paramount [23].
Research Reagent Solutions
Table 3. Essential Reagents for PCR Optimization Experiments
| Reagent/Category | Specific Function | Application Notes |
|---|---|---|
| Magnesium Chloride (MgCl₂) | DNA polymerase cofactor; stabilizes primer-template binding | Concentration typically 1.5-3.0 mM; requires template-specific optimization [5] |
| High-Fidelity Polymerase | DNA synthesis with proofreading capability | EVO HS, Q5, Phusion systems; offer 50-100X greater accuracy than Taq [45] |
| dNTPs | Nucleotide substrates for DNA synthesis | Standard concentration 200µM; quality affects fidelity [45] |
| PCR Enhancers | Modify template secondary structure | DMSO, betaine, formamide for GC-rich templates [44] |
| Hot-Start Mechanisms | Inhibit polymerase at room temperature | Antibody-based, chemical modification, or physical separation methods [46] |
| Buffer Systems | Maintain optimal pH and ionic strength | Often contain Tris-HCl, (NH₄)₂SO₄, and stabilizers [44] |
Mg²⁺ concentration represents a fundamental parameter in PCR optimization, with precise modulation necessary for achieving application-specific performance requirements. The established optimal range of 1.5–3.0 mM serves as a valuable starting point, but template complexity, polymerase characteristics, and application requirements necessitate empirical determination of ideal concentrations for each experimental system [5]. For high-fidelity applications such as cloning and sequencing, lower Mg²⁺ concentrations (1.5–2.0 mM) generally favor accuracy, while diagnostic applications targeting low-copy templates may benefit from slightly higher concentrations (2.0–3.5 mM) to enhance sensitivity [45] [21]. Continued advancements in enzyme engineering, including the development of novel DNA polymerase variants with expanded functionality, promise to further simplify reaction optimization while maintaining the critical balance between efficiency, specificity, and fidelity across diverse PCR applications [23].
Troubleshooting Common PCR Pitfalls Through Magnesium Concentration Adjustment
In the precise world of polymerase chain reaction (PCR) optimization, magnesium ions (Mg²⁺) are not merely a buffer component but a fundamental cofactor governing enzymatic activity and reaction fidelity. A deficiency in Mg²⁺ concentration directly halts the polymerization process, leading to the complete absence of amplification—a critical failure mode in sensitive applications from diagnostic testing to cloning [47] [1]. The Mg²⁺ ion is an essential cofactor for all thermostable DNA polymerases, where it directly facilitates the catalytic incorporation of nucleotides into the growing DNA strand [2] [1]. Beyond catalysis, Mg²⁺ stabilizes the double-stranded primer-template hybrid by neutralizing the negative charges on the phosphate backbones of DNA strands, thereby promoting successful annealing [47]. Operating outside the optimal Mg²⁺ window, particularly at excessively low concentrations, inactivates the DNA polymerase and results in a total failure of the assay, presenting a clear risk to data integrity in pharmaceutical and research settings [47] [48].
Quantitative Data: Establishing the Optimal Magnesium Window
Empirical and meta-analytical data consistently define a specific concentration range for magnesium chloride (MgCl₂) that supports efficient amplification while minimizing errors. Deviations from this range produce predictable, quantifiable pathologies in the PCR outcome.
Table 1: The Effect of MgCl₂ Concentration on PCR Performance and Fidelity
| MgCl₂ Concentration | Amplification Yield | Specificity & Fidelity | Observed Experimental Artifacts |
|---|---|---|---|
| < 1.0 mM | Very low to none [1] | Not applicable (no product) | Primer binding fails; polymerase is inactive, leading to complete amplification failure and no visible band on a gel [47] [1]. |
| 1.5 - 3.0 mM (Optimal Range) | High [5] [20] | High [47] | Defined, single band of the expected amplicon size. A 2025 meta-analysis identified this as the optimal range, with a logarithmic relationship to DNA melting temperature (every 0.5 mM increase raises Tm by ~1.2°C) [5] [20]. |
| > 3.0 mM | High, but with spurious products [47] | Low [47] [48] | Multiple bands or smearing on a gel due to non-specific primer binding and misincorporation; reduced polymerase fidelity [47] [49]. |
The required Mg²⁺ concentration is not universal but is influenced by template complexity. Genomic DNA templates typically require higher concentrations within the optimal range compared to more straightforward templates like plasmid DNA or cDNA [5] [20]. Furthermore, the concentration of dNTPs must be considered, as Mg²⁺ binds to dNTPs in the reaction mix. An excess of dNTPs can chelate the available Mg²⁺, effectively creating a Mg²⁺-deficient environment even if the initial MgCl₂ concentration was nominally correct [47] [2].
Experimental Protocols: Methodologies for Systematic Mg²⁺ Optimization
Magnesium Titration as a Standard Optimization Procedure
To empirically determine the optimal Mg²⁺ concentration for a specific assay, a titration experiment is the most reliable approach. The following protocol is adapted from common practices detailed across multiple sources [47] [49] [48].
- Prepare a Master Mix: Create a standard master mix containing all PCR components except for MgCl₂. This includes buffer (without magnesium), DNA template, primers, dNTPs, and DNA polymerase.
- Aliquot the Reaction: Dispense equal volumes of the master mix into multiple PCR tubes or wells.
- Titrate MgCl₂: Add MgCl₂ stock solution to each aliquot to create a concentration gradient. A typical range is 1.0 mM to 4.0 mM, in increments of 0.5 mM [49] [48].
- Run PCR: Place the reactions in a thermal cycler and initiate the standard PCR program.
- Analyze Results: Resolve the PCR products using agarose gel electrophoresis. The well with the strongest, single, correct band and the least background smearing indicates the optimal MgCl₂ concentration for that assay.
Protocol for Overcoming Calcium-Induced Inhibition
In forensic and ancient DNA analysis, calcium (Ca²⁺) from bone samples can be a potent PCR inhibitor by competitively binding the polymerase's active site in place of Mg²⁺ [50]. The following protocol demonstrates a method to reverse this inhibition.
- Sample Setup: Set up parallel PCR reactions containing a known inhibitor (e.g., calcium ions) and a fixed, typically optimal, concentration of MgCl₂.
- Intervention: To the test reaction, add the calcium chelator ethylene glycol-bis(2-aminoethylether)-N,N,N′,N′-tetraacetic acid (EGTA) at a final concentration of 0.5-1.0 mM [50]. A control reaction should contain no EGTA.
- Amplification and Quantification: Perform qPCR amplification. The results will show recovery of amplification efficiency in the EGTA-treated sample, as evidenced by a lower Cq value compared to the inhibited control, demonstrating that EGTA chelates Ca²⁺, freeing the Mg²⁺ to act as the correct cofactor [50].
The Scientist's Toolkit: Essential Reagents for PCR Fidelity Research
Table 2: Key Research Reagents for Investigating Mg²⁺ Effects in PCR
| Reagent / Tool | Function in Research | Key Consideration |
|---|---|---|
| High-Fidelity Polymerase Mix (e.g., Q5, KOD) | Engineered for high accuracy; KOD polymerase shows higher resistance to metal ion inhibition in comparative studies [47] [50]. | Essential for studies where product sequence integrity is paramount, such as cloning or sequencing. |
| MgCl₂ Stock Solution (1M, molecular biology grade) | Allows for precise molarity adjustments in reaction optimization via titration [47]. | Solution must be fully thawed and vortexed before use to avoid concentration gradients from freeze-thaw cycles [48]. |
| PCR Additives (e.g., DMSO, Betaine) | Assist in amplifying difficult templates (e.g., GC-rich regions) by destabilizing secondary structures, which can change Mg²⁺ requirements [47] [49] [48]. | DMSO can inhibit Taq polymerase; concentration must be optimized (typically 2-10%) [47] [48]. |
| Chelating Agents (e.g., EGTA, EDTA) | EGTA is used to selectively chelate Ca²⁺ to alleviate its inhibition [50]. EDTA is a general chelator that can be used to create defined, low-Mg²⁺ conditions. | Carry-over EDTA from DNA extraction kits can sequester Mg²⁺ and cause amplification failure [47]. |
| Gradient Thermal Cycler | Enables parallel testing of a range of annealing temperatures and/or Mg²⁺ concentrations in a single run, drastically speeding up optimization [47]. | Critical for high-throughput and systematic optimization workflows. |
Signaling Pathways and Workflows: A Mg²⁺-Centric View of PCR
The following diagram synthesizes the core biochemical role of Mg²⁺ in the PCR extension step and the logical decision pathway for troubleshooting failures related to low Mg²⁺.
Figure 1: Mg²⁺ Function in PCR and Failure Diagnosis. The top section illustrates the critical, dual role of Mg²⁺ as an enzyme cofactor and a binding partner for dNTPs during the catalytic formation of a new phosphodiester bond. The bottom section provides a logical workflow for diagnosing and resolving the specific symptom of no amplification, tracing it back to low Mg²⁺ and its direct biochemical consequences.
In the context of evaluating PCR fidelity, the magnesium ion (Mg²⁺) concentration emerges as one of the most crucial parameters influencing reaction specificity. As an essential cofactor for DNA polymerase activity, Mg²⁺ facilitates the catalytic incorporation of nucleotides and stabilizes the interaction between primers and template DNA [1] [2]. However, exceeding the optimal concentration range initiates a cascade of molecular events that compromise amplification fidelity, primarily manifesting as non-specific bands and smearing on agarose gels. This systematic analysis compares the effects of excessively high Mg²⁺ concentrations against optimal and deficient conditions, providing quantitative data and experimental methodologies for researchers seeking to optimize PCR protocols for diagnostic and research applications. Understanding these concentration-dependent effects is fundamental to designing reliable genetic assays, particularly when working with complex templates or demanding applications such as drug development research.
Mechanistic Insights: How Mg²⁺ Concentration Governs PCR Specificity
The Dual Role of Mg²⁺ in PCR Biochemistry
Magnesium chloride (MgCl₂) serves two fundamental biochemical functions in PCR amplification. Primarily, it acts as an essential cofactor for thermostable DNA polymerases (e.g., Taq polymerase) by facilitating the formation of phosphodiester bonds during DNA synthesis [1]. The Mg²⁺ ion binds to the dNTP at its alpha phosphate group, enabling the removal of beta and gamma phosphates and allowing the resulting dNMP to form a phosphodiester bond with the 3' OH group of the adjacent nucleotide [1]. Secondarily, Mg²⁺ stabilizes the DNA duplex by binding to negatively charged phosphate groups along the DNA backbone, thereby reducing electrostatic repulsion between primer and template strands [1] [2]. This binding effectively increases the melting temperature (Tm) of primer-template hybrids, with meta-analyses demonstrating a logarithmic relationship where every 0.5 mM increase in MgCl₂ within the 1.5-3.0 mM range raises the melting temperature by approximately 1.2°C [20] [5].
Concentration-Dependent Effects on Reaction Fidelity
The fidelity of PCR amplification exhibits a strong dependence on Mg²⁺ concentration, with distinct performance phases observed across the concentration spectrum:
- Deficient Range (<1.5 mM): Inadequate Mg²⁺ availability limits DNA polymerase activity, resulting in weak amplification or complete PCR failure due to impaired primer extension and insufficient enzyme catalysis [6] [1].
- Optimal Range (1.5-4.5 mM): This zone supports balanced reaction kinetics where primers anneal specifically to target sequences with high fidelity, producing strong, specific amplicons without secondary products [6].
- Excessive Range (>4.5 mM): Supra-optimal concentrations reduce primer annealing stringency through increased Tm and stabilization of mismatched hybrids, promoting non-specific binding and erroneous amplification [6] [1] [51].
The following diagram illustrates the logical relationship between Mg²⁺ concentration and PCR outcomes:
Comparative Performance Analysis: Quantitative Effects of Mg²⁺ Concentration
Systematic Comparison of PCR Outcomes Across Mg²⁺ Concentrations
The following table synthesizes quantitative data from multiple studies investigating Mg²⁺ effects on PCR performance, particularly focusing on template specificity and amplification efficiency:
Table 1: Comparative Analysis of PCR Performance Across Mg²⁺ Concentration Ranges
| Mg²⁺ Concentration | Amplification Efficiency | Band Specificity | Common Artifacts | Recommended Applications |
|---|---|---|---|---|
| Low (≤1.5 mM) | Weak to non-detectable [6] [1] | N/A | No amplification or faint target band | Not recommended for standard applications |
| Optimal (1.5-4.5 mM) | Strong, reproducible amplification [6] | High specificity, single clear band [6] | Minimal non-specific products | Standard PCR, diagnostic applications, cloning |
| High (≥4.5 mM) | High but non-specific [6] [1] | Low specificity, multiple bands [6] [1] | Non-specific bands, smearing, primer dimers [6] [1] | Specialized applications requiring reduced stringency |
Template-Specific Magnesium Optimization Requirements
Template characteristics significantly influence optimal Mg²⁺ requirements. The following table summarizes evidence-based recommendations for different template types:
Table 2: Template-Specific Mg²⁺ Optimization Guidelines Based on Experimental Data
| Template Type | Recommended Mg²⁺ Range | Experimental Evidence | Special Considerations |
|---|---|---|---|
| Standard Templates | 1.5-2.0 mM [1] | Routine optimization sufficient for most applications | Balance specificity and yield |
| High GC Content | 2.0-4.5 mM [20] | Higher concentrations improve melting of secondary structures | May require additives (DMSO, betaine) [30] |
| Genomic DNA | 2.0-4.0 mM [5] | Meta-analysis shows genomic DNA requires higher concentrations than plasmid DNA | Compensate for complexity and potential inhibitors |
| Complex Mixtures (e.g., fecal metagenomes) | Up to 7.0 mM [51] | Study detected optimal tet genes at 7 mM with specific primer set | Concentration depends on primer degeneracy and sample inhibitors |
Experimental Protocols: Methodologies for Mg²⁺ Optimization
Standard Mg²⁺ Titration Protocol
To systematically optimize Mg²⁺ concentration for any PCR application, the following methodology adapted from foundational protocols is recommended [30]:
- Reaction Setup: Prepare a master mix containing all standard PCR components except MgCl₂. Aliquot equal volumes into thin-walled PCR tubes.
- Mg²⁺ Titration: Add MgCl₂ to achieve final concentrations spanning 1.0 mM to 5.0 mM in 0.5 mM increments. Include a negative control without template DNA.
- Thermal Cycling: Perform amplification using predetermined cycling parameters with an annealing temperature approximately 5°C below the calculated primer Tm.
- Product Analysis: Separate PCR products by agarose gel electrophoresis. Identify the concentration yielding the strongest specific amplification with minimal background.
This protocol enables researchers to empirically determine the optimal Mg²⁺ concentration for their specific template-primer system, balancing the competing demands of specificity and efficiency.
Factorial Experimental Design for Challenging Templates
For complex applications such as amplifying antibiotic resistance genes from microbiome samples, a more comprehensive approach is warranted. A published investigation into tet gene detection employed a 4 × 3 factorial design testing four MgCl₂ concentrations (1.75 mM, 3 mM, 5 mM, and 7 mM) against three different PCR cycle numbers (25, 30, and 35 cycles) [51]. This methodology revealed that both parameters significantly influenced the apparent diversity and relative distribution of resistance genes detected. The researchers subsequently cloned and sequenced products from each condition, demonstrating that optimal detection of ribosomal protection protein tet genes in swine feces occurred at 35 cycles with 7 mM MgCl₂ [51]. This factorial approach provides a robust framework for optimizing challenging PCR applications where template complexity or primer degeneracy complicates standard optimization.
The Scientist's Toolkit: Essential Reagents for PCR Optimization
Table 3: Key Research Reagent Solutions for PCR Optimization Experiments
| Reagent | Function in PCR | Optimization Considerations |
|---|---|---|
| MgCl₂ | DNA polymerase cofactor; stabilizes primer-template binding [1] [2] | Critical optimization parameter (1.5-4.5 mM standard range); concentration affects specificity and yield [6] |
| DNA Polymerase | Enzymatic synthesis of new DNA strands [2] | 1-2 units/50 µL reaction standard; may increase with inhibitors; excess can cause non-specific products [2] |
| Primers | Sequence-specific amplification initiation [30] [2] | 0.1-1 µM final concentration; excessive primers promote mispriming; design with 40-60% GC content [30] |
| dNTPs | Nucleotide substrates for DNA synthesis [2] | 0.2 mM each dNTP standard; balance with Mg²⁺ concentration (Mg²⁺ binds dNTPs) [2] |
| Template DNA | Target sequence for amplification [2] | 1-1000 ng genomic DNA; excess causes non-specific amplification; too little reduces yield [30] [2] |
| PCR Additives (DMSO, BSA, Betaine) | Modifiers of nucleic acid hybridization [30] | DMSO (1-10%) improves GC-rich amplification; BSA (10-100 µg/mL) counteracts inhibitors [30] |
Discussion: Integrated Optimization Strategy for PCR Fidelity
The empirical evidence consistently demonstrates that excessively high Mg²⁺ concentrations fundamentally alter PCR thermodynamics, reducing the stringency of primer annealing and permitting amplification at non-target sites [6] [1] [51]. This mechanistic understanding explains the observed correlation between elevated Mg²⁺ and the appearance of multiple non-specific bands and smearing on analytical gels. Beyond Mg²⁺ concentration, contemporary meta-analyses emphasize the importance of template-specific optimization, with genomic DNA and GC-rich templates typically requiring higher concentrations than simple plasmid templates [20] [5].
Successful PCR optimization requires a systematic approach that considers the interplay between all reaction components. While Mg²⁺ concentration represents a pivotal parameter, its effects are modulated by primer design, dNTP concentration, template quality, and thermal cycling conditions. Researchers should implement iterative optimization protocols, beginning with Mg²⁺ titration followed by adjustment of annealing temperatures and cycle numbers. For particularly challenging applications, such as amplifying diverse gene families from complex metagenomic samples, factorial experimental designs may be necessary to reveal condition-dependent amplification biases [51]. This comprehensive approach to PCR optimization ensures maximum fidelity and reproducibility, essential requirements for both basic research and drug development applications.
In polymerase chain reaction (PCR) optimization, achieving high yield is a fundamental objective that often hinges on the precise modulation of reaction components. Among these, magnesium ions (Mg²⁺) and deoxynucleoside triphosphates (dNTPs) form a critical biochemical partnership that directly governs polymerase activity, reaction efficiency, and ultimately, amplification success. Mg²⁺ serves as an essential cofactor for DNA polymerase, enabling the enzyme to catalyze the formation of phosphodiester bonds between nucleotides [2]. Simultaneously, the availability of dNTPs—the building blocks of DNA synthesis—must be carefully balanced against Mg²⁺ concentration, as these components exist in a tightly regulated equilibrium.
The relationship between Mg²⁺ and dNTPs is particularly crucial when addressing the common PCR symptom of low yield. Mg²⁺ directly stabilizes the primer-template complex and facilitates enzyme function, while also binding to dNTPs to form the actual substrates incorporated by the polymerase [2] [47]. This binding creates a competitive dynamic where elevated dNTP concentrations can effectively chelate available Mg²⁺, reducing the free Mg²⁺ pool necessary for polymerase activity and potentially leading to insufficient amplification [2]. Understanding this interdependence provides researchers with a powerful leverage point for systematically troubleshooting and optimizing PCR protocols to maximize product yield across diverse applications and template types.
Comparative Analysis: Experimental Data on Mg²⁺ and dNTP Concentrations
Magnesium Concentration Effects on PCR Performance
Table 1: Experimental Effects of Mg²⁺ Concentration on PCR Performance
| Mg²⁺ Concentration | Amplification Efficiency | Specificity | Recommended Application Context |
|---|---|---|---|
| < 1.5 mM | Significantly reduced or absent | High (if any product) | Suboptimal for most PCR applications |
| 1.5–2.0 mM | Optimal | High | Standard PCR with Taq polymerase [52] |
| 2.0–3.0 mM | High | Moderate to high | Genomic DNA templates [5] |
| 3.0–4.0 mM | High | Reduced | Problematic templates (high GC content) |
| > 4.0 mM | Variable | Significantly reduced | Generally not recommended due to spurious products |
A comprehensive meta-analysis of 61 peer-reviewed studies established a strong logarithmic relationship between MgCl₂ concentration and DNA melting temperature, with optimal amplification ranges typically falling between 1.5 and 3.0 mM [5]. Within this range, every 0.5 mM increase in MgCl₂ was associated with a 1.2 °C increase in melting temperature, directly influencing primer-template binding stability [5]. The analysis further revealed that template complexity significantly affects optimal Mg²⁺ requirements, with genomic DNA templates generally requiring higher concentrations (closer to 3.0 mM) compared to more straightforward plasmid DNA templates [5] [2].
Experimental evidence demonstrates that deviation from optimal Mg²⁺ concentrations produces characteristic symptoms. When Mg²⁺ is too low, no PCR product is typically observed due to insufficient polymerase activity [52]. Conversely, excessively high Mg²⁺ concentrations (typically >3-4 mM) promote non-specific amplification through reduced enzymatic fidelity and increased tolerance of mismatched primer-template hybrids [47] [52]. This manifests as multiple bands or smearing on agarose gels, complicating result interpretation and reducing target product yield.
dNTP Concentration and Balancing with Mg²⁺
Table 2: Effects of dNTP Concentration on PCR Outcomes
| dNTP Concentration (each) | Amplification Yield | Fidelity | Mg²⁺ Requirement |
|---|---|---|---|
| 50–100 µM | Reduced | Higher | Lower (1.5–2.0 mM) |
| 200 µM (standard) | High | Standard | 1.5–2.0 mM [52] |
| 300–400 µM | Higher (for long amplicons) | Reduced | Higher (2.5–3.5 mM) |
| > 400 µM | Potentially inhibited | Significantly reduced | Significantly higher |
The standard recommended concentration for each dNTP in PCR is typically 200 µM, which provides optimal balance between yield and fidelity for most applications [52]. At this concentration, sufficient nucleotide building blocks are available for efficient amplification without introducing excessive errors. However, research indicates that lower dNTP concentrations (50-100 µM) can enhance fidelity by improving the discrimination capability of DNA polymerase, though this comes at the cost of reduced overall yield [52]. Conversely, higher dNTP concentrations (300-400 µM) may improve yields for longer amplicons but generally reduce fidelity and require proportional increases in Mg²⁺ concentration [52].
The critical relationship between Mg²⁺ and dNTPs cannot be overstated. Mg²⁺ binds to dNTPs to form the actual substrates that DNA polymerase recognizes and incorporates [2]. Consequently, the molar concentration of Mg²⁺ must exceed the total dNTP concentration to maintain an adequate pool of free Mg²⁺ for polymerase function. The total dNTP concentration in a standard PCR reaction (4 nucleotides × 200 µM each = 800 µM) requires careful balancing with Mg²⁺ levels, which is why most protocols recommend 1.5-2.0 mM Mg²⁺ [2] [52]. When dNTP concentrations are increased, Mg²⁺ must be supplemented proportionally to compensate for this chelation effect.
Methodological Framework: Optimization Protocols
Systematic Optimization of Mg²⁺ Concentration
Based on experimental data from multiple sources, the following systematic approach is recommended for optimizing Mg²⁺ concentration to address low yield:
Initial Setup: Begin with a standard concentration of 1.5 mM Mg²⁺ and prepare a series of reactions with increments of 0.5 mM, typically ranging from 1.0 mM to 4.0 mM [52]. These reactions should maintain consistent concentrations of all other components, particularly dNTPs.
Template-Specific Adjustments: For genomic DNA templates, focus testing in the 2.0-3.0 mM range, as the meta-analysis indicates these complex templates often require higher Mg²⁺ concentrations [5]. For plasmid or cDNA templates, the 1.5-2.5 mM range is typically more appropriate [2].
Analysis and Refinement: Evaluate amplification results using agarose gel electrophoresis. Identify the concentration that produces the highest yield of the specific target product with minimal non-specific amplification. If necessary, perform a finer titration (0.25 mM increments) around the most promising concentration from the initial test.
The optimization workflow for Mg²⁺ follows a logical progression from initial setup to refined testing, with decision points at each stage based on experimental outcomes. This workflow can be visualized as follows:
Complementary dNTP Titration Approach
While Mg²⁺ optimization often resolves yield issues, simultaneous consideration of dNTP concentrations may be necessary:
Standard Protocol: Begin with 200 µM of each dNTP and 1.5-2.0 mM Mg²⁺ as a baseline [52].
Fidelity-Focused Adjustment: For applications requiring high fidelity (e.g., cloning, sequencing), reduce dNTP concentrations to 50-100 µM each and proportionally lower Mg²⁺ to 1.0-1.5 mM [52].
Yield-Focused Adjustment: For challenging templates where yield is prioritized over fidelity, increase dNTPs to 300-400 µM each and supplement Mg²⁺ to 2.5-3.5 mM to maintain free Mg²⁺ availability [2].
Special Applications: When using modified nucleotides (dUTP, biotin-dUTP, etc.), verify polymerase compatibility and consider that incorporation efficiency may differ from standard dNTPs, potentially requiring adjusted concentrations and cycling conditions [2].
Research Reagent Solutions for Optimization
Table 3: Essential Research Reagents for PCR Optimization
| Reagent Category | Specific Examples | Function in Optimization | Usage Notes |
|---|---|---|---|
| Magnesium Salts | MgCl₂, MgSO₄ | Essential cofactor for DNA polymerase; stabilizes primer-template binding | Concentration significantly impacts specificity and yield [5] [52] |
| dNTP Mixtures | dATP, dCTP, dGTP, dTTP | Building blocks for DNA synthesis; concentration affects fidelity and Mg²⁺ availability | Typically used at 200 µM each; balanced concentrations critical [2] [52] |
| DNA Polymerases | Taq, Pfu, KOD, engineered variants | Catalyzes DNA synthesis; different enzymes have varying Mg²⁺ and dNTP requirements | Taq optimal for yield; proofreading enzymes for fidelity [47] [23] |
| Buffer Systems | Tris-HCl, (NH₄)₂SO₄, KCl | Maintain pH and ionic strength; some contain proprietary enhancers | Can affect Mg²⁺ availability; use matching manufacturer buffer [47] |
| Template DNA | Genomic DNA, cDNA, plasmid DNA | Target source for amplification; quality and complexity affect component requirements | 1pg-10ng plasmid DNA; 1ng-1µg genomic DNA recommended [2] [52] |
| Additives | DMSO, betaine, formamide | Reduce secondary structure; modify effective melting temperatures | DMSO (0-10%) lowers Tm; use with GC-rich templates [47] [53] |
Discussion: Integrated Troubleshooting for Low Yield
Template-Dependent Optimization Strategies
The optimization of Mg²⁺ and dNTP concentrations cannot be divorced from template characteristics, as different template types present unique challenges. Genomic DNA templates generally require higher Mg²⁺ concentrations (2.0-3.0 mM) compared to plasmid DNA due to their complexity and potential co-purification of inhibitors [5] [2]. For GC-rich templates, which often exhibit strong secondary structure, the addition of enhancers like DMSO (2-10%) or betaine (1-2 M) can significantly improve yield, but these additives may necessitate adjustment of Mg²⁺ concentrations and annealing temperatures [47] [53]. When working with low-copy number targets, increasing template amount (within recommended limits) and using higher fidelity conditions (lower dNTPs) may provide better results than simply increasing Mg²⁺ concentrations [2] [52].
Recent advances in polymerase engineering have created enzymes with modified cofactor requirements that can simplify optimization challenges. Novel Taq polymerase variants have been developed with enhanced reverse transcriptase activity, reducing dependency on specific Mg²⁺ concentrations for efficient cDNA synthesis and amplification [23]. Similarly, specialized polymerases for long-range PCR or high-GC content templates often include optimized buffer systems that minimize the need for extensive Mg²⁺ titration [47].
Addressing Common Optimization Artifacts
Several characteristic symptoms during PCR optimization can be directly traced to Mg²⁺ and dNTP imbalances:
No Amplification: This result frequently indicates insufficient Mg²⁺ concentration (typically <1.0-1.5 mM) or excessive dNTPs that chelate available Mg²⁺ [52]. Resolution involves increasing Mg²⁺ concentration while maintaining standard dNTP levels, or reducing dNTP concentrations if they are elevated.
Non-specific Bands/Smearing: This common artifact typically results from excessive Mg²⁺ concentration (>3.0-4.0 mM) or insufficiently stringent annealing conditions [47] [52]. Systematic reduction of Mg²⁺ in 0.5 mM increments combined with increased annealing temperature often resolves this issue.
Primer-Dimer Formation: This phenomenon occurs when primers self-anneal rather than binding to the template, often exacerbated by excessive primer concentrations, insufficient template, or suboptimal Mg²⁺ levels [2]. Reducing primer concentration to 0.1-0.5 µM and optimizing Mg²⁺ typically reduces primer-dimer formation.
The interplay between Mg²⁺, dNTPs, and annealing temperature creates a three-way optimization landscape where adjustment of one parameter may necessitate refinement of others. For example, increasing Mg²⁺ concentration to improve yield may allow for increased annealing temperature to maintain specificity, creating an optimized balance tailored to specific template-primer systems [47] [53].
The fine-tuning of Mg²⁺ and dNTP concentrations represents a fundamental strategy for addressing the common PCR symptom of low yield. Evidence from multiple studies confirms that the optimal Mg²⁺ concentration typically falls between 1.5 and 3.0 mM, while standard dNTP concentrations of 200 µM for each nucleotide provide the best balance between yield and fidelity for most applications [5] [52]. The critical interdependence between these components necessitates coordinated optimization, as dNTPs effectively reduce available Mg²⁺ through chelation, directly impacting polymerase activity.
A systematic approach to optimization should prioritize initial Mg²⁺ titration using 0.5 mM increments across the 1.0-4.0 mM range, with template-specific focus areas guided by template complexity [5] [52]. Subsequent refinement should address dNTP concentrations based on application priorities—favoring lower concentrations (50-100 µM) for high-fidelity applications and potentially higher concentrations (300-400 µM) for challenging templates where yield is paramount [52]. This methodological framework, combined with an understanding of template-specific requirements and polymerase characteristics, provides researchers with a powerful systematic approach to resolving low yield in PCR applications across diverse research contexts.
In the pursuit of PCR fidelity, the meticulous optimization of magnesium ion (Mg²⁺) concentration is a cornerstone of robust assay design. However, the effective availability of this essential cofactor is critically dependent on a frequently overlooked variable: template purity. PCR inhibitors present in impure nucleic acid preparations can systematically chelate Mg²⁺, disrupt enzyme activity, and quench fluorescent signals, leading to inaccurate quantification and failed reactions. This guide objectively examines how common inhibitors interfere with Mg²⁺ availability and compares strategic solutions—from sample purification to reagent enhancement—enabling researchers to maintain quantification accuracy across challenging sample types.
The Biochemical Interplay: Inhibitors and Mg²⁺ Chelation
Mg²⁺ is an indispensable cofactor for DNA polymerase activity, stabilizing the enzyme-DNA complex and facilitating primer annealing [30]. Its effective concentration in the reaction mix dictates PCR efficiency and specificity. However, various inhibitory substances commonly found in biological and environmental samples can sequester Mg²⁺, rendering it unavailable for the polymerization reaction.
The mechanisms of PCR inhibition extend beyond simple Mg²⁺ chelation. As illustrated below, inhibitors act through multiple biochemical pathways that ultimately compromise PCR efficiency and data reliability.
Common inhibitors and their specific effects on PCR are detailed in the table below.
Table 1: Common PCR Inhibitors and Their Effects on Mg²⁺ Availability and PCR Efficiency
| Inhibitor Source | Example Inhibitors | Primary Mechanism of Interference | Impact on Effective Mg²⁺ |
|---|---|---|---|
| Biological Samples | Hemoglobin, lactoferrin, immunoglobulins [54] [55] | Enzyme inhibition; Mg²⁺ chelation [55] | Reduced availability |
| Environmental Samples | Humic acids, fulvic acids, tannins [56] [54] | Interact with nucleic acids/polymerase; Mg²⁺ chelation [54] | Significant sequestration |
| Laboratory Reagents | Phenol, SDS, high salt concentrations [55] [57] | Disrupt primer binding; precipitate template [55] | Alters reaction equilibrium |
Comparative Analysis of Correction Strategies
Researchers can employ multiple strategies to overcome inhibition and restore Mg²⁺ availability. The optimal approach often depends on the source and severity of inhibition, as well as downstream application requirements.
Sample Purification Techniques
Enhanced nucleic acid purification is the first line of defense against PCR inhibitors. The table below compares the most common purification methods.
Table 2: Comparison of PCR Product Purification Techniques for Inhibitor Removal
| Technique | Mechanism | Effectiveness Against Inhibitors | Drawbacks | Best For |
|---|---|---|---|---|
| Spin Column [58] [57] | Silica-membrane binding in presence of chaotropic salts [58] | Good for salts, proteins, enzymes [58] | Less effective on small fragments; plastic waste [57] | Routine purification; high-quality DNA [57] |
| Magnetic Beads [57] | Magnetic beads with DNA-binding coatings [57] | High yield and purity; scalable [57] | Higher setup costs; optimization complexity [57] | High-throughput labs; automation [57] |
| Enzymatic Clean-Up [57] | Degrades primers/dNTPs with enzymes [57] | Moderate (does not remove all contaminants) [57] | Incomplete inhibitor removal [57] | High-throughput; cost-sensitive labs [57] |
PCR Enhancers and Additives
Chemical additives can be incorporated into the PCR master mix to neutralize inhibitors and stabilize reaction components. Their effectiveness varies based on the inhibitor type.
Table 3: Efficacy of Common PCR Enhancers Against Various Inhibitors
| Enhancer | Recommended Concentration | Proposed Mechanism | Documented Efficacy |
|---|---|---|---|
| Bovine Serum Albumin (BSA) | 10-100 μg/mL [30] | Binds to inhibitors like humic acids and polyphenolics [56] | Reduced failure rate to 0.1% in buccal swab samples [59] |
| T4 Gene 32 Protein (gp32) | Varies | Binds single-stranded DNA, stabilizes denaturation [56] | Counteracts humic acid inhibition [56] |
| Dimethyl Sulfoxide (DMSO) | 1-10% [30] | Lowers DNA melting temperature [56] | Variable; can inhibit some polymerases at high concentrations [56] |
| Betaine | 0.5 M to 2.5 M [30] | Equalizes strand stability, especially in GC-rich templates [30] | Stabilizes polymerase; improves amplification of complex templates [30] |
Inhibitor-Tolerant Polymerase Blends
Choosing a DNA polymerase engineered for inhibitor tolerance provides a straightforward solution. These specialized enzymes maintain activity even when Mg²⁺ availability is suboptimal or in the presence of common inhibitors.
Experimental Data: One study demonstrated that the inhibitor-tolerant Phusion Flash polymerase enabled successful direct PCR from blood samples, reducing DNA profiling time from 10-12 hours to 2-3 hours by eliminating extensive purification [54]. Similarly, Promega's GoTaq Endure qPCR Master Mix is specifically formulated for reliable amplification of challenging samples like blood, soil, and plant-derived nucleic acids [55].
Decision Framework for Researchers
Selecting the most appropriate strategy depends on the specific inhibition context. The workflow below outlines a systematic approach to diagnosing and correcting for PCR inhibitors.
The Scientist's Toolkit: Essential Research Reagent Solutions
The following reagents and kits represent key solutions for managing Mg²⁺ availability in the presence of inhibitors.
Table 4: Essential Research Reagents for Overcoming PCR Inhibition
| Reagent/Kits | Primary Function | Key Advantage | Reference |
|---|---|---|---|
| Monarch Spin PCR & DNA Cleanup Kit | Purifies DNA fragments (50 bp to 25 kb) | High recovery with low elution volumes; no pH monitoring | [60] |
| GoTaq Endure qPCR Master Mix | Inhibitor-tolerant qPCR reactions | Consistent amplification in blood, soil, plant samples | [55] |
| PureLink Quick Gel Extraction & PCR Purification Combo Kit | Dual-purpose gel extraction and PCR cleanup | Rapid purification (<30 min) for sequencing/cloning | [58] |
| Bovine Serum Albumin (BSA) | Additive to bind inhibitors in reaction mix | Cost-effective; proven efficacy in buccal swabs | [59] |
| microCLEAN PCR Cleanup Reagent | Single-tube PCR purification | Reduces plastic waste; no harmful chemicals | [57] |
Template purity is a fundamental determinant of effective Mg²⁺ availability in PCR, directly impacting assay fidelity and quantification accuracy. While common inhibitors systematically chelate this essential cofactor, researchers have multiple validated strategies at their disposal. Data demonstrates that combining rigorous purification protocols—such as magnetic bead-based cleanup—with reagent-level enhancements like BSA or inhibitor-tolerant polymerases provides the most robust correction. The optimal approach depends on the specific inhibitor profile, but the systematic framework presented here enables informed decision-making for reliable PCR outcomes across diverse sample types.
The pursuit of high-fidelity amplification, especially within the critical context of evaluating polymerase chain reaction (PCR) fidelity across different magnesium concentrations, demands a sophisticated understanding of buffer chemistry. The interplay between magnesium ions (Mg²⁺) and chemical additives like DMSO and betaine is not merely additive but often synergistic, creating a reaction environment that can overcome some of the most persistent challenges in molecular biology, such as amplifying GC-rich templates and complex secondary structures. While Mg²⁺ acts as an essential cofactor for DNA polymerase activity, its concentration directly influences enzyme specificity and fidelity [44] [61]. Conversely, additives like DMSO and betaine function as isostabilizing agents, modulating the physical properties of the DNA template to facilitate strand separation and primer annealing [62] [63]. This guide provides a detailed, objective comparison of how integrating Mg²⁺ optimization with these additives enhances PCR performance, supported by experimental data and tailored for applications in genetic research and diagnostic assay development.
Core Concepts: Mg²⁺ and Additives in PCR
Magnesium Ions (Mg²⁺): The Essential Cofactor
Magnesium chloride (MgCl₂) is a non-negotiable component of any PCR buffer. Its primary role is to serve as a cofactor for DNA polymerase, enabling the enzyme to catalyze the formation of phosphodiester bonds between nucleotides [44]. The Mg²⁺ ions facilitate the binding of the polymerase to the DNA template and stabilize the interaction with incoming dNTPs. However, the concentration is a critical variable that requires precise optimization. Too little Mg²⁺ results in low enzyme activity and poor yield, while an excess can reduce specificity by stabilizing non-specific primer-template interactions and increasing error rates [44] [61]. This narrow optimal range makes Mg²⁺ titration a fundamental first step in any PCR fidelity study.
Structure-Disrupting Additives: DMSO and Betaine
GC-rich DNA templates pose a significant challenge due to their propensity to form stable secondary structures, such as hairpins and G-quadruplexes, which resist denaturation and can cause polymerase stalling [44] [62]. DMSO and betaine are among the most effective additives for countering these effects.
DMSO (Dimethyl Sulfoxide): This reagent acts by disrupting the hydrogen bonding and base stacking interactions that stabilize DNA secondary structures. By doing so, it effectively lowers the melting temperature (Tₘ) of the DNA, making it easier to denature complex templates during the PCR heating steps [62] [64]. Typical working concentrations range from 1% to 10%, with 5% often being a standard starting point for optimization [65].
Betaine (Trimethylglycine): Betaine is an amino acid derivative that functions as a isostabilizing agent. It equilibrates the differential melting temperatures between AT and GC base pairs by preferentially hydrating AT-rich regions and destabilizing GC-rich regions. This action helps to "relax" the entire DNA molecule, making GC-rich areas more accessible to the polymerase and preventing the enzyme from stalling at secondary structures [63] [62]. Common effective concentrations used in PCR are between 0.5 M and 1.5 M [63] [61].
Table 1: Characteristics of Key PCR Additives
| Additive | Primary Mechanism of Action | Typical Working Concentration | Main Application |
|---|---|---|---|
| DMSO | Disrupts hydrogen bonding, lowers DNA Tₘ | 5% (1-10% range) [65] | GC-rich templates, reduces secondary structures [62] |
| Betaine | Equalizes Tₘ of AT and GC base pairs | 0.5 M - 1.5 M [63] | GC-rich templates, prevents polymerase stalling [62] |
| Proline | Combined with betaine for synergistic effect | 0.4 M [63] | Enhances specificity for high-GC, complex structures [63] |
Comparative Performance Analysis
Synergistic Effects of Combined Optimization
While both Mg²⁺ and additives can improve PCR individually, their combined and optimized use often yields the most significant gains, particularly for difficult templates. The buffer system in PCR Kit V2 exemplifies this synergy, employing a dual-ion system where Mg²⁺ activates the polymerase and ammonium ions from ammonium sulfate enhance primer specificity by disrupting weak hydrogen bonds in mismatched base pairs [44]. Introducing betaine and DMSO into this optimized buffer provides an additional layer of control over template secondary structures, which is crucial for targets with GC content exceeding 65% [44].
Experimental data demonstrates that a specialized 2x GC-rich master mix containing 3 mM MgCl₂, 0.5 M betaine, and 0.4 M proline (an additive similar in function to DMSO) successfully amplified DNA fragments with GC content ranging from 61.5% to 74.8%, which failed to amplify in standard buffers [63]. This combination assists in template denaturation and improves primer binding specificity, thereby aiding DNA polymerase extension through problematic regions.
Table 2: PCR Success Rate with Standard vs. Optimized Buffer Systems
| Template Complexity | Standard Buffer Success Rate | Optimized Buffer with Additives Success Rate |
|---|---|---|
| GC-rich (>70%) | 50–60% | >90% [44] |
| GC-rich (ITS2 plant barcode) | 42% | 91.6% with 5% DMSO; 100% with DMSO/betaine substitution strategy [65] |
| Long DNA (>5 kb) | Low stability | High stability [44] |
| Multiplex Compatibility | Moderate | Excellent [44] |
Impact on Fidelity and Specificity
The ultimate goal of combining these components is to achieve high-fidelity amplification with minimal off-target products. Elevated Mg²⁺ concentrations are a known source of reduced fidelity, as they can promote non-specific binding and increase mis-incorporation rates [61]. The use of structure-disrupting additives like DMSO and betaine can indirectly improve fidelity by allowing the reaction to run efficiently at a lower, more specific Mg²⁺ concentration. Furthermore, by preventing polymerase stalling at secondary structures, these additives reduce the likelihood of the enzyme introducing errors [62].
For the highest fidelity requirements, such as in next-generation sequencing (NGS) library preparation or cloning, high-fidelity polymerases with proofreading activity (3'→5' exonuclease) are recommended. Enzymes like the EVO HS Super-Fidelity DNA Polymerase boast ~50–100X the fidelity of conventional Taq polymerase and are also compatible with DMSO or betaine for GC-rich templates, demonstrating that additive optimization is applicable across a range of enzyme types [45].
Experimental Protocols and Workflows
A Standardized Workflow for PCR Optimization
Adopting a systematic approach is crucial for successfully optimizing PCR conditions for challenging templates. The following workflow, derived from established methodologies, ensures that variables are tested in a logical sequence.
Detailed Protocol: Optimizing a GC-Rich PCR
This protocol is adapted from studies that successfully amplified extremely GC-rich constructs using a combination of additives and optimized magnesium [63] [62].
1. Reagent Setup: Prepare a master mix on ice with the following components for a 50 µL reaction:
- 1X EVO Buffer (or other high-fidelity polymerase buffer) [45]
- 200 µM dNTPs (each) [45]
- 0.5 µM forward and reverse primers [45]
- 1–2 U of a high-fidelity DNA polymerase (e.g., EVO HS, Phusion) [45]
- Template DNA: 10 pg to 500 ng, quantity requires optimization [45]
- Nuclease-free water to final volume
2. Magnesium and Additive Titration: Aliquot the master mix and create reaction tubes with:
- Mg²⁺ Gradient: A range of 1.5 mM to 2.5 mM (or up to 4.0 mM for some polymerases) is a suitable starting point [61] [45].
- Additives:
3. Thermal Cycling: Use a thermal cycler with a gradient function for the annealing step.
- Initial Denaturation: 98°C for 30 sec [45]
- Denaturation (30-35 cycles): 98°C for 5-10 sec [45]
- Annealing: Use a temperature gradient (e.g., 55–72°C) to identify the optimal temperature simultaneously [64]. A Touchdown PCR protocol, starting 5-10°C above the calculated Tₘ and decreasing 0.5-1°C per cycle for the first 10-15 cycles, is highly effective for improving specificity [64].
- Extension: 72°C at 15-30 sec/kb [45]
- Final Extension: 72°C for 5 min [45]
4. Analysis: Analyze PCR products by agarose gel electrophoresis. The optimal condition is the one that yields a single, bright band of the expected size.
The Scientist's Toolkit: Essential Research Reagent Solutions
A successful optimization experiment relies on a core set of reliable reagents and instruments. The following table details key materials and their functions.
Table 3: Essential Reagents and Tools for PCR Optimization
| Item / Reagent Solution | Function / Application Note |
|---|---|
| High-Fidelity DNA Polymerase (e.g., EVO HS, Q5, Phusion) | Provides high accuracy and robust amplification of complex templates; often includes a hot-start mechanism [45]. |
| PCR Kit V2-type Buffer | A pre-optimized system containing Tris-HCl (pH buffer), (NH₄)₂SO₄ (primer specificity), and often a balanced Mg²⁺ concentration [44]. |
| Molecular Biology Grade DMSO | A structure-disrupting additive for lowering DNA Tₘ; use high-purity grade to avoid contaminants [62]. |
| Betaine (Trimethylglycine) | An isostabilizing agent for equalizing Tₘ across DNA regions; typically prepared as a 5M stock solution [63]. |
| L-Proline | An additive that, in combination with betaine, shows synergistic effects in amplifying high-GC and complex structure templates [63]. |
| Gradient Thermal Cycler | Essential instrumentation for efficiently testing different annealing and denaturation temperatures in a single run [64]. |
The strategic combination of Mg²⁺ optimization with structure-disrupting additives represents a powerful and often essential approach for advanced PCR applications. As the field moves towards more complex genetic analyses, including the detection of multi-gene panels and low-frequency variants, the demand for robust, first-attempt amplification will only increase. Future developments will likely see the emergence of novel polymerases, such as the engineered Taq variants with innate reverse transcriptase activity for one-tube RT-PCR [12], which will also benefit from these refined buffer chemistries. Furthermore, the integration of machine learning in primer design and predictive modeling of buffer compatibility promises to reduce the empirical optimization burden on researchers. For now, a methodical understanding of the synergistic relationship between Mg²⁺, DMSO, and betaine provides a reliable foundation for overcoming the most stubborn PCR challenges, thereby ensuring fidelity and reproducibility in genetic research and molecular diagnostics.
Validation and Comparative Analysis of PCR Fidelity Across Magnesium Conditions
The fidelity of a DNA polymerase is defined as the accuracy with which it copies a DNA template sequence, a critical parameter for experiments whose outcomes depend on the correct DNA sequence, such as cloning, single nucleotide polymorphism (SNP) analysis, and next-generation sequencing (NGS) applications [66]. Fidelity comparisons are expressed either in absolute terms, such as the number of errors per thousand nucleotides, or in relative terms by using Taq DNA polymerase as a reference standard (e.g., 1X for Taq) [66]. The consequences of low fidelity are false variant calls in sequencing and mutated clones, making fidelity assessment a cornerstone of rigorous molecular biology.
The biochemical basis of fidelity revolves around two primary mechanisms: nucleotide selection and proofreading. The polymerase active site preferentially selects and incorporates correct nucleotides based on Watson-Crick base pairing; an incorrect nucleotide leads to a suboptimal architecture, slowing incorporation and allowing for dissociation [66]. Furthermore, many high-fidelity polymerases possess a 3´→5´ exonuclease (proofreading) domain. This domain detects and excises misincorporated nucleotides from the growing DNA strand before permanent incorporation, providing a significant reduction in error rates [66] [47]. Factors such as buffer composition, deoxynucleoside triphosphate (dNTP) concentrations, and particularly the concentration of magnesium ions (Mg2+) can profoundly influence these mechanisms and the resulting fidelity [2] [67].
Methodologies for Assessing Polymerase Fidelity
The evolution of fidelity assessment methods mirrors advances in sequencing technology, progressing from indirect phenotypic assays to direct, high-throughput sequencing capable of detecting errors with single-base resolution.
Classical Methods: LacZ-Based Assays
The pioneering work in fidelity measurement utilized the lacZα gene in M13 bacteriophage [66]. Errors during DNA synthesis in a specific region of the β-galactosidase gene lead to a loss of function, which is detected by a color change in bacterial colonies on agar plates containing X-gal (from blue to white). A modification of this assay, the Barnes assay, amplified the entire lacZ gene and portions of drug resistance genes via PCR, followed by cloning, transformation, and blue/white colony screening [66].
- Workflow: PCR amplification (16 cycles) → ligation and cloning → transformation → blue/white colony counting.
- Output: Mutation frequency derived from the ratio of white (mutant) to blue (functional) colonies.
- Advantages: Relatively high-throughput for its time and cost-effective.
- Limitations: Indirect measurement; only detects errors in specific regions that affect the phenotype (e.g., only 349 of 1,900 bases in lacZ cause a color change); cannot identify the type or location of specific mutations [66].
Sanger Sequencing-Based Fidelity Assays
The advent of Sanger sequencing offered a more direct readout of polymerase errors. This method involves amplifying a target gene, cloning the products, and then sequencing individual clones to identify all mutations within the amplicon [66].
- Workflow: PCR amplification (e.g., 25 cycles) → cloning → Sanger sequencing of individual clones.
- Output: Direct sequence data allowing identification of substitution rates, insertions, and deletions (indels).
- Advantages: Direct detection of all mutation types within the sequenced region.
- Limitations: Lower throughput due to cost and labor; the number of nucleotides sequenced is often too low to accurately quantify the error rate of ultra-high-fidelity polymerases with statistical confidence [66]. For example, sequencing 440,000 nucleotides for Q5 polymerase identified only two errors, suggesting an error rate of ~1 in a million, but this lacks precision [66].
Next-Generation Sequencing (NGS)-Based Fidelity Assays
NGS platforms overcome the throughput limitations of previous methods, enabling the sequencing of millions to billions of nucleotides and providing statistically robust error rate measurements, even for the most accurate polymerases [68] [66].
- Barcoded (Illumina) Sequencing: This approach uses primers containing unique molecular identifiers (barcodes) to tag individual template molecules [68]. After PCR and sequencing, reads sharing the same barcode are grouped. A mutation present in all reads of a group is identified as an error originating from the initial reverse transcription or cDNA synthesis step, while random errors are attributed to PCR or the sequencing process itself [68]. A reported limitation is that its lower threshold for accurate error rate determination is around 1 × 10⁻⁶ errors per base, which is near the intrinsic error rate of some high-fidelity enzymes [66].
- Single-Molecule Real-Time (SMRT) Sequencing: PacBio SMRT sequencing allows for the direct sequencing of PCR products without an intermediary cloning or amplification step [66]. It achieves high accuracy by repeatedly sequencing the same molecule to generate a highly accurate consensus sequence. The background error rate for this method is exceptionally low (9.6 × 10⁻⁸ errors per base), making it suitable for quantifying the fidelity of proofreading polymerases with high precision [66]. This method can also capture a wider range of replication errors, including template switching and sequence recombination.
The following workflow summarizes the key steps in a barcoded NGS fidelity assay, as used for evaluating reverse transcriptases [68]:
Comparative Analysis of Fidelity Measurement Methods
Table 1: Comparison of Polymerase Fidelity Assessment Methodologies
| Method | Key Principle | Detection Limit | Throughput | Key Advantage | Key Disadvantage |
|---|---|---|---|---|---|
| LacZ (Barnes Assay) | Phenotypic color selection in E. coli | Not precisely defined | Moderate | Historically high-throughput, cost-effective | Indirect; only detects mutations in specific codons |
| Sanger Sequencing | Direct sequencing of cloned PCR products | ~1 x 10⁻⁵ errors/base [66] | Low | Direct readout of all mutation types in the amplicon | Low throughput limits statistical power for high-fidelity enzymes |
| Barcoded NGS | Sequencing with unique molecular identifiers | ~1 x 10⁻⁶ errors/base [66] | Very High | Can distinguish errors from different reaction steps (RT, PCR) [68] | Background error rate limits quantification of ultra-high-fidelity polymerases |
| SMRT Sequencing | Direct sequencing and circular consensus | ~9.6 x 10⁻⁸ errors/base [66] | High | Lowest background; captures full spectrum of error types [66] | Higher cost per sample compared to short-read NGS |
The Critical Role of Magnesium Concentration in PCR Fidelity
Among various reaction components, magnesium chloride (MgCl₂) concentration is one of the most crucial parameters affecting PCR success, specificity, and critically, fidelity [20] [47] [2]. Mg²⁺ ions act as an essential cofactor for DNA polymerase activity, stabilizing the enzyme's active structure and facilitating the binding of its actual substrate, the dNTP-Mg²⁺ complex [2].
Mechanisms of Magnesium Action
Mg²⁺ influences fidelity through multiple interconnected mechanisms:
- Enzyme Cofactor: It is directly required for the polymerase to catalyze the nucleophilic attack and incorporate dNTPs into the growing DNA chain [2].
- Nucleotide Binding: Mg²⁺ forms a soluble complex with dNTPs, creating the substrate recognized by the polymerase. The concentration of free Mg²⁺ is therefore critical and is determined by the concentrations of dNTPs, which chelate the ion, and other chelators like EDTA [67].
- Reaction Stringency: Mg²⁺ stabilizes the double-stranded DNA formed during primer annealing. Its concentration directly affects the melting temperature (Tm) of the primer-template hybrid; a meta-analysis demonstrated a logarithmic relationship between MgCl₂ concentration and DNA melting temperature, with Tm consistently rising by approximately 0.5–0.7°C for every 0.5 mM increment in MgCl₂ within the 1.5–3.0 mM range [20].
- Fidelity and Specificity: Optimal Mg²⁺ concentration is a balance. Low Mg²⁺ reduces enzyme activity and can lead to poor yield, while excess Mg²⁺ promotes non-specific amplification and lowers fidelity by reducing the polymerase's specificity for correct base pairing [47] [67].
Experimental Optimization of Magnesium Concentration
Optimizing Mg²⁺ is essential for any fidelity-related research. The typical optimal concentration ranges from 1.5 to 2.5 mM, but this must be determined empirically for each primer-template system [47]. The relationship between Mg²⁺ concentration and PCR performance can be visualized as having distinct functional phases [20]:
The most effective method for optimization is a Mg²⁺ titration experiment, typically across a range of 0.5 to 5.0 mM in 0.5 mM increments, while keeping all other parameters constant [47]. The ideal concentration is identified as the one that produces the highest yield of the specific target amplicon with minimal to no non-specific products. It is crucial to note that any change in dNTP concentration necessitates re-optimization of Mg²⁺, as dNTPs chelate Mg²⁺ ions [2].
Experimental Data and Comparative Performance
Fidelity Measurements of Commercial Polymerases
Modern fidelity assays using NGS provide precise error rates for a wide range of commercially available DNA polymerases. The following table synthesizes data from PacBio SMRT sequencing, which offers a low background and high statistical power [66].
Table 2: Polymerase Fidelity Measurements via SMRT Sequencing [66]
| DNA Polymerase | Substitution Rate (per base per doubling) | Accuracy (1/Substitution Rate) | Fidelity Relative to Taq | Key Feature |
|---|---|---|---|---|
| Q5 High-Fidelity | 5.3 × 10⁻⁷ | 1,887,000 | 280X | Ultra-high fidelity, proofreading |
| Phusion | 3.9 × 10⁻⁶ | 256,000 | 39X | Proofreading |
| Deep Vent | 4.0 × 10⁻⁶ | 250,000 | 44X | Proofreading |
| Pfu | 5.1 × 10⁻⁶ | 196,000 | 30X | Proofreading |
| PrimeSTAR GXL | 8.4 × 10⁻⁶ | 119,000 | 18X | Proofreading |
| KOD | 1.2 × 10⁻⁵ | 83,000 | 12X | Proofreading |
| Kapa HiFi | 1.6 × 10⁻⁵ | 63,000 | 9.4X | Proofreading |
| Taq | 1.5 × 10⁻⁴ | 6,700 | 1X (Reference) | Non-proofreading |
| Deep Vent (exo-) | 5.0 × 10⁻⁴ | 2,000 | 0.3X | Proofreading-deficient |
The data clearly demonstrates the profound impact of proofreading activity. Comparing Deep Vent (exo+) to its exonuclease-deficient variant (Deep Vent exo-) shows that the proofreading domain provides a 125-fold decrease in error rate [66]. Furthermore, engineered polymerases like Q5 set the benchmark for ultra-high-fidelity amplification.
Reverse Transcriptase Fidelity Assessment
The barcoded NGS method has been effectively applied to evaluate the fidelity of reverse transcriptases (RTs). A study using this approach reported the following error rates:
- MMLV RT (MM4 variant): 0.75-1.0 × 10⁻⁴ errors/base
- RTX (DNA polymerase with RT activity): 0.75-1.0 × 10⁻⁴ errors/base
- Wild-type HIV-1 RT: 2.6 × 10⁻⁴ errors/base [68]
This highlights that fidelity varies significantly among RTs and that the NGS-based method can precisely evaluate them under different reaction conditions in a high-throughput manner [68].
Essential Reagents and Protocols for Fidelity Research
The Scientist's Toolkit for Fidelity Assessment
Table 3: Key Research Reagent Solutions for Fidelity Experiments
| Reagent / Material | Function in Fidelity Assessment | Key Considerations |
|---|---|---|
| High-Fidelity DNA Polymerase | Amplifies target with minimal errors for downstream analysis. | Select based on proofreading activity and documented error rate (e.g., Q5, Pfu) [66] [47]. |
| Ultra-Pure dNTPs | Balanced building blocks for accurate DNA synthesis. | Use equimolar, high-quality mixes to prevent incorporation errors; concentration affects Mg²⁺ availability [2] [67]. |
| Molecular Biology Grade MgCl₂ | Essential polymerase cofactor and reaction stabilizer. | Concentration must be optimized; purity is critical to avoid contaminants that affect reaction kinetics [20] [2]. |
| Optimized Reaction Buffer | Provides optimal pH and ionic environment for the polymerase. | Often supplied with the enzyme; may require supplementation or adjustment based on Mg²⁺ titration results. |
| Barcoded Primers (for NGS) | Tags individual template molecules to trace error origins. | Essential for NGS-based fidelity assays to distinguish errors from different reaction steps [68]. |
| Standardized Control Template | Provides a consistent, high-quality sequence for error rate calculation. | Plasmid DNA or synthetic oligonucleotides with a known sequence are ideal [66]. |
| NGS Library Prep Kit | Prepares the amplified DNA for high-throughput sequencing. | Should be selected for low bias and high efficiency to avoid introducing artifacts. |
Detailed Experimental Protocol: Magnesium Titration for Fidelity Optimization
This protocol provides a step-by-step guide for optimizing Mg²⁺ concentration to achieve high fidelity in PCR.
Objective: To determine the optimal MgCl₂ concentration for a specific primer-template pair that maximizes yield and specificity, thereby creating conditions for high-fidelity amplification.
Materials:
- Target DNA template (e.g., 10-50 ng genomic DNA)
- Forward and reverse primers (0.1-1 μM final concentration)
- High-fidelity DNA polymerase with corresponding reaction buffer (without MgCl₂)
- 25 mM MgCl₂ stock solution
- PCR-grade dNTP mix (e.g., 10 mM each)
- Nuclease-free water
- Thermal cycler
Procedure:
- Prepare Master Mix: Create a master mix for n+1 reactions (where n is the number of Mg²⁺ conditions) containing nuclease-free water, reaction buffer, primers, dNTPs, DNA polymerase, and the DNA template. Mix thoroughly by gentle pipetting.
- Aliquot and Titrate Mg²⁺: Aliquot equal volumes of the master mix into 8 PCR tubes. Add the 25 mM MgCl₂ stock to each tube to create a final concentration series (e.g., 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, and 4.0 mM). Include a negative control (no template DNA).
- Thermal Cycling: Run the PCR using the following generalized cycling conditions, adjusting the annealing temperature (Ta) as required for your primers:
- Initial Denaturation: 98°C for 30 seconds
- 30 cycles of:
- Denaturation: 98°C for 10 seconds
- Annealing: (Ta)°C for 20 seconds
- Extension: 72°C for 15-30 seconds per kb
- Final Extension: 72°C for 2 minutes
- Hold: 4°C
- Analysis: Analyze 5-10 μL of each reaction product by agarose gel electrophoresis. Visualize the gel under UV light.
- Interpretation: Identify the Mg²⁺ concentration that yields the strongest, most specific band of the expected size with the least background smearing or non-specific bands. This concentration represents the optimal balance between activity and specificity for your system and should be used for all subsequent high-fidelity applications with this primer-template set.
The methodologies for assessing DNA polymerase fidelity have evolved dramatically, from phenotypic screens in bacteria to highly precise NGS-based analyses capable of detecting single-base errors in millions of nucleotides. This progression has revealed vast differences in performance among enzymes, with proofreading polymerases like Q5 offering a 280-fold increase in fidelity over standard Taq polymerase [66]. Within this framework, the optimization of reaction components, particularly MgCl₂ concentration, remains a foundational and non-negotiable step. The interplay between Mg²⁺, enzyme activity, and reaction stringency means that empirical titration is essential for establishing conditions that support maximum fidelity. As molecular techniques demand ever-greater accuracy, the continued refinement of these assessment and optimization strategies will be paramount for ensuring the validity of genetic data in research, diagnostics, and therapeutic development.
In polymerase chain reaction (PCR) experiments, magnesium ions (Mg²⁺) function as an essential cofactor for all thermostable DNA polymerases. The concentration of Mg²⁺ directly influences three critical aspects of the reaction: enzyme activity, primer-template annealing stability, and crucially, the fidelity of the amplification process [47]. Maintaining the integrity of the DNA sequence during amplification is paramount for applications whose outcomes depend on correct sequence data, such as cloning, single-nucleotide polymorphism (SNP) analysis, and next-generation sequencing (NGS) [69]. This guide objectively compares the performance of PCR assays under low, optimal, and high Mg²⁺ conditions, providing quantitative data on error rates and practical protocols for researchers aiming to optimize reaction fidelity.
The fidelity of DNA replication describes the accuracy with which a DNA polymerase copies a template sequence. This accuracy is maintained through multiple mechanisms, including the polymerase's inherent ability to select the correct nucleoside triphosphate and its capacity to proofread and excise misincorporated nucleotides via a 3'→5' exonuclease domain present in some high-fidelity enzymes [69]. The concentration of Mg²⁺ is a key variable that influences these processes, making its optimization a critical step in experimental design.
Quantitative Data on Error Rates and Mg²⁺ Concentration
Fidelity Comparison of DNA Polymerases
The error rate of a DNA polymerase is a foundational metric for understanding fidelity. Different polymerases exhibit vastly different intrinsic error rates, which can be further influenced by reaction conditions like Mg²⁺ concentration. The table below summarizes the documented error rates for common polymerases, providing a baseline for fidelity comparisons.
Table 1: DNA Polymerase Fidelity Measurements Based on PacBio SMRT Sequencing [69]
| DNA Polymerase | Substitution Rate (per base per doubling) | Accuracy (1/Substitution Rate) | Fidelity Relative to Taq |
|---|---|---|---|
| Q5 High-Fidelity | 5.3 × 10⁻⁷ | 1,870,763 | 280X |
| Phusion | 3.9 × 10⁻⁶ | 255,118 | 39X |
| Pfu | 5.1 × 10⁻⁶ | 195,275 | 30X |
| Deep Vent | 4.0 × 10⁻⁶ | 251,129 | 44X |
| Taq | 1.5 × 10⁻⁴ | 6,456 | 1X |
| Deep Vent (exo-) | 5.0 × 10⁻⁴ | 2,020 | 0.3X |
The Direct Impact of Mg²⁺ Concentration on PCR Performance
The Mg²⁺ concentration must be carefully titrated for any given PCR system. Its effect is not linear, and deviations from the optimal range can lead to severe performance issues, impacting both yield and accuracy.
Table 2: Impact of Mg²⁺ Concentration on PCR Specificity and Fidelity [47]
| Mg²⁺ Condition | Typical Concentration Range | Effect on Enzyme Activity | Effect on Specificity & Fidelity | Overall Effect on Reaction Yield |
|---|---|---|---|---|
| Low Mg²⁺ | < 1.5 mM | Reduced polymerase activity, leading to inefficient primer extension and low product yield. | Increased specificity in some cases, but overall process is inefficient. | Significantly reduced or complete amplification failure. |
| Optimal Mg²⁺ | 1.5 - 3.0 mM (often requires titration) | Maximal enzyme activity and processivity. | High fidelity and specificity; correct base pairing is favored. | Maximum yield of the desired specific product. |
| High Mg²⁺ | > 3.0 mM | Slightly increased non-beneficial activity. | Markedly reduced fidelity and specificity; promotes non-specific amplification and misincorporation. | High yield of non-specific products and smeared bands; desired product may be obscured. |
The mechanism behind this fidelity loss is rooted in the molecular role of Mg²⁺. Structural studies of DNA polymerases reveal that a catalytic Mg²⁺ ion (Metal A) coordinates the 3’-OH of the primer terminus, positioning it for an in-line nucleophilic attack on the alpha-phosphate of the incoming nucleotide [70]. Improper Mg²⁺ concentrations distort this active site geometry, compromising the enzyme's ability to discriminate against incorrect nucleotides and leading to the error rates observed in practical experiments.
Experimental Protocols for Quantifying PCR Errors
Accurately determining polymerase error rates requires sophisticated assays that move beyond simple gel electrophoresis. Below are detailed methodologies for two common approaches used in rigorous fidelity testing.
Next-Generation Sequencing (NGS) Based Fidelity Assay
This protocol leverages the high throughput of NGS to directly sequence PCR products and identify misincorporations.
- Step 1: Template and Amplification. A plasmid DNA template (e.g., LacZ gene) that is virtually devoid of nucleotide errors is selected. The target amplicon is amplified for a limited number of cycles (e.g., 16-25 cycles) using the polymerase and the specific Mg²⁺ conditions under investigation [69].
- Step 2: Library Preparation and Sequencing. The PCR products are prepared for sequencing. To account for sequencing errors, some protocols use molecular barcoding or single-molecule, real-time (SMRT) sequencing, which generates a highly accurate consensus sequence for each read without the need for an intermediary amplification step that could introduce its own errors [69].
- Step 3: Data Analysis. The sequenced reads are aligned to the reference template sequence. The error rate is calculated as the number of observed substitutions, insertions, and deletions (indels) per base per doubling event, factoring in the number of effective amplification cycles [69].
Functional LacZα Complementation Assay
This classical genetic assay provides a functional readout of mutation frequency.
- Step 1: Amplification and Cloning. A plasmid containing the LacZα gene is amplified via PCR using the test conditions. The resulting PCR products are ligated into an appropriate vector and transformed into an E. coli host strain [69].
- Step 2: Phenotypic Screening. Transformed bacteria are plated on agar containing the substrate X-gal. Colonies with a functional LacZα gene (blue) indicate error-free amplification, while colonies with a mutated, non-functional gene (white) indicate a PCR error was introduced.
- Step 3: Error Rate Calculation. The number of white and blue colonies is counted. The error rate is inferred from the frequency of white colonies after correcting for background mutations and error propagation during PCR [69]. This method, while high-throughput, only detects errors that inactivate the gene.
The Scientist's Toolkit: Essential Reagents for Fidelity Research
Table 3: Key Research Reagent Solutions for PCR Fidelity Experiments
| Reagent / Material | Function in Experiment | Key Considerations |
|---|---|---|
| High-Fidelity DNA Polymerase (e.g., Q5, Pfu) | Catalyzes DNA synthesis with high accuracy due to proofreading activity (3'→5' exonuclease). | Provides a low-error baseline; ~10-280x more accurate than standard Taq [69]. |
| Standard Taq Polymerase | Catalyzes DNA synthesis without proofreading activity. | Serves as a fidelity control; error rate ~1 error per 6,500 bases [69]. |
| MgCl₂ or MgSO₄ Solution | Source of essential Mg²⁺ cofactor. | Concentration must be titrated (e.g., 0.5 mM steps from 1.0 to 3.5 mM) for optimization [47]. |
| dNTP Mix | Provides nucleotide substrates for DNA synthesis. | Imbalanced dNTP concentrations can increase error rates; use high-purity stocks. |
| Control Plasmid DNA (e.g., LacZ) | A well-characterized, error-free template for fidelity assays. | Allows for direct quantification of errors introduced during PCR [69]. |
| Buffer Additives (e.g., DMSO, Betaine) | Aids in amplifying complex templates and can influence fidelity. | DMSO lowers Tm and can improve specificity but must be accounted for in Tm calculations [53]. |
Visualizing the Relationship Between Mg²⁺ and PCR Fidelity
The following diagram illustrates the mechanistic role of Mg²⁺ in the polymerase active site and the consequences of its concentration on PCR outcomes.
Diagram Title: Mg²⁺ Role and Impact on PCR
The quantitative data and experimental protocols presented in this guide underscore a critical, non-linear relationship between Mg²⁺ concentration and PCR fidelity. An optimal Mg²⁺ concentration (typically 1.5-3.0 mM) is fundamental for achieving a balance between high yield and high accuracy, as it supports the correct geometry in the polymerase active site necessary for faithful DNA replication [70]. Straying from this optimum in either direction is detrimental: low Mg²⁺ cripples polymerase activity, while high Mg²⁺ systematically reduces fidelity, promoting misincorporation and non-specific amplification that can compromise experimental results [47].
For researchers in drug development and diagnostics, where sequence integrity is paramount, these findings highlight the necessity of empirical Mg²⁺ optimization for every new set of primers and template. Relying on standard buffer formulations is insufficient. A systematic titration of Mg²⁺, combined with the use of a high-fidelity proofreading polymerase for sensitive applications, provides the most robust strategy for ensuring data reliability and reproducibility in molecular biology workflows.
The polymerase chain reaction (PCR) serves as a foundational technique in molecular biology, enabling the amplification and analysis of genetic material across diverse research and diagnostic applications. Within this process, magnesium chloride (MgCl₂) acts as a critical cofactor for DNA polymerase activity, influencing enzyme fidelity, processivity, and primer-template binding specificity. The concentration of Mg²⁺ presents a particular challenge in the detection and diversity analysis of multi-gene families, where sequence homology can lead to cross-hybridization and amplification bias. This case study objectively compares the performance of PCR under different magnesium concentrations, providing supporting experimental data to guide researchers in optimizing protocols for complex genetic targets. Within the broader context of evaluating PCR fidelity, understanding the nuanced role of Mg²⁺ becomes paramount for generating reliable, reproducible results in gene family studies, mutation detection, and phylogenetic analyses.
The Dual Role of Mg²⁺ in PCR and Primer Specificity
Magnesium ions serve as an essential catalytic cofactor for DNA polymerase function, facilitating the nucleotidyl transferase reaction during DNA synthesis. Beyond this fundamental role, Mg²⁺ concentration directly impacts primer-template binding stability and specificity by reducing electrostatic repulsion between the primer and the negatively charged DNA backbone. This dual function creates a narrow optimal concentration range that must be empirically determined for different primer-template systems.
In the context of multi-gene families characterized by high sequence homology, suboptimal Mg²⁺ concentrations can dramatically alter amplification efficiency and product distribution. Excessive magnesium (typically >4.5 mM for many systems) reduces reaction stringency, permitting non-specific priming and cross-amplification of paralogous gene family members. This can result in heterogeneous amplicon pools that complicate downstream interpretation. Conversely, inadequate magnesium (<1.0 mM) diminishes polymerase processivity and reduces overall yield, potentially failing to detect low-abundance family members and introducing representation bias in diversity assessments [71].
The influence of Mg²⁺ extends to polymerase fidelity, with elevated concentrations correlating with increased misincorporation rates. This effect manifests particularly in mononucleotide and dinucleotide repeat regions, where Mg²⁺ concentrations above 2.0 mM can exacerbate polymerase slippage, generating artificial "shadow bands" and length polymorphisms that mimic genuine genetic diversity [72]. These artifacts present significant challenges when analyzing microsatellite markers within gene families or conducting mutation screening in repetitive genomic regions.
Experimental Protocols for Mg²⁺ Optimization
Establishing a Magnesium Concentration Gradient
A standardized approach to determining optimal Mg²⁺ concentration involves creating a titration series while holding all other reaction components constant. The following protocol adapts methodologies from published optimization procedures [71] [21]:
Reagents and Equipment:
- 10X PCR buffer (without MgCl₂)
- 25 mM MgCl₂ stock solution
- Template DNA (10-50 ng/μL)
- Forward and reverse primers (10 μM each)
- dNTP mix (10 mM each)
- DNA polymerase (1-2.5 U/μL)
- Thermocycler
- Gel electrophoresis system
Procedure:
- Prepare a master mix containing all reaction components except MgCl₂ and template DNA.
- Aliquot equal volumes of the master mix into 8 PCR tubes.
- Add MgCl₂ stock solution to achieve final concentrations of: 1.0 mM, 1.5 mM, 2.0 mM, 2.5 mM, 3.0 mM, 3.5 mM, 4.0 mM, and 4.5 mM.
- Add template DNA to each tube and initiate thermal cycling using predetermined parameters.
- Analyze products via agarose gel electrophoresis with ethidium bromide staining.
- Document band intensity, specificity, and product size distribution for each concentration.
Evaluating Multi-Gene Family Amplification
To assess Mg²⁺ effects on multi-gene family diversity, researchers can employ a model system with known paralogous sequences. The following protocol utilizes the CYP51C gene family in Fusarium species as described in recent methodological comparisons [21]:
Primer Design:
- Target conserved regions flanking variable domains within the gene family
- Design primers with balanced GC content (40-60%)
- Verify specificity in silico against all known family members
Amplification and Analysis:
- Perform parallel amplifications across the Mg²⁺ gradient established in section 3.1.
- Clone products from each concentration into sequencing vectors.
- Sequence multiple clones (minimum 20 per concentration) to assess representation of different family members.
- Quantify amplification bias by comparing the relative frequency of each paralog across Mg²⁺ concentrations.
Table 1: Effect of Mg²⁺ Concentration on PCR Specificity and Yield
| Mg²⁺ Concentration (mM) | Band Specificity | Relative Yield | Non-specific Products | Recommended Application |
|---|---|---|---|---|
| 1.0 | Low | 25% | None | Limited utility |
| 1.5 | High | 65% | None | High-specificity applications |
| 2.0 | High | 100% | None | Standard amplification |
| 2.5 | Moderate | 115% | Minimal | Complex templates |
| 3.0 | Moderate | 120% | Moderate | Low-copy targets |
| 3.5 | Low | 110% | Significant | Not recommended |
| 4.0 | Low | 95% | Extensive | Not recommended |
| 4.5 | Very low | 80% | Extensive | Not recommended |
Comparative Performance Data
Magnesium-Dependent Fidelity Assessment
Error rate comparisons across polymerase types reveal significant Mg²⁺ dependence in replication fidelity. As shown in Table 2, conventional Taq polymerase demonstrates substantially higher error rates compared to proofreading enzymes, with both groups exhibiting Mg²⁺-sensitive mutation frequencies. Notably, mononucleotide and dinucleotide repeats display particular vulnerability to Mg²⁺-dependent slippage, with error rates increasing approximately 3-fold between 1.5 mM and 4.5 mM Mg²⁺ concentrations [15] [72].
Table 2: Polymerase Fidelity Comparison Under Standard Mg²⁺ Conditions (2.0 mM)
| DNA Polymerase | Error Rate (errors/bp/duplication) | Fidelity Relative to Taq | Optimal Mg²⁺ Range (mM) |
|---|---|---|---|
| Taq | 1-20 × 10⁻⁵ | 1x | 1.5-2.5 |
| AccuPrime-Taq High Fidelity | ~1.0 × 10⁻⁵ | ~9x better | 1.5-2.0 |
| KOD Hot Start | ~7 × 10⁻⁷ | ~28x better | 1.0-2.0 |
| Pfu | 1-2 × 10⁻⁶ | 6-10x better | 2.0-3.0 |
| Phusion Hot Start | 4-9.5 × 10⁻⁷ | 24-50x better | 1.5-2.0 |
| Pwo | ~1 × 10⁻⁶ | ~10x better | 2.0-3.0 |
The fidelity differential between polymerase families has profound implications for multi-gene family analysis. In a direct sequencing study of 94 unique DNA targets, Taq polymerase introduced mutations in approximately 3.0-5.6 × 10⁻⁵ bases sequenced, while high-fidelity enzymes like Pfu and Phusion exhibited error rates below 1 × 10⁻⁶ [15]. These mutations can create artificial diversity in gene family analyses, particularly when assessing single-nucleotide polymorphisms among paralogs.
Impact on Multi-Gene Family Detection
Recent methodological comparisons demonstrate that Mg²⁺ optimization critically influences detection capabilities for complex gene families. In developing detection assays for Fusarium tricinctum, researchers found that Mg²⁺ concentrations between 2.0-4.5 mM yielded markedly different sensitivity thresholds across three molecular detection platforms [21]:
Quantitative PCR: Maximum sensitivity (3.1 fg/μL detection limit) occurred at 2.0 mM Mg²⁺, with a 10-fold reduction in sensitivity at 4.5 mM due to non-specific amplification.
Nested PCR: Optimal performance required 2.5 mM Mg²⁺ in the primary reaction and 2.0 mM in the secondary reaction, balancing yield with specificity across two amplification rounds.
LAMP: This isothermal method functioned across a broader Mg²⁺ range (2.0-8.0 mM), with peak performance at 6.0 mM, highlighting technique-specific cofactor requirements.
These findings underscore the necessity of technique-specific Mg²⁺ optimization, particularly when targeting multi-gene families where sequence conservation might promote cross-amplification at elevated Mg²⁺ concentrations.
Visualization of Mg²⁺ Effects on PCR Specificity
The following diagram illustrates the mechanism by which Mg²⁺ concentration influences amplification specificity and fidelity in multi-gene family analysis:
Advanced Applications and Novel Polymerase Engineering
Recent advancements in polymerase engineering offer promising solutions to Mg²⁺-associated challenges in multi-gene family analysis. Novel Taq polymerase variants with enhanced reverse transcriptase activity demonstrate exceptional thermostability and maintain functionality across broad Mg²⁺ ranges (1.5-4.5 mM), enabling simultaneous reverse transcription and DNA amplification without requiring viral reverse transcriptases [12]. These engineered enzymes facilitate multiplex detection of various RNA targets in a single reaction, significantly simplifying complex gene expression analyses across gene families.
The development of novel detection platforms like NAPTUNE (Nucleic acids and Protein Biomarkers Testing via Ultra-sensitive Nucleases Escalation) demonstrates alternative approaches that minimize amplification-related bias in multi-analyte detection [73]. This system employs apurinic/apyrimidinic endonuclease 1 (APE1) and Pyrococcus furiosus Argonaute (PfAgo) in a tandem cascade, achieving femtomolar to attomolar sensitivity without target amplification, thereby bypassing Mg²⁺-dependent fidelity issues entirely.
Similarly, RNA aptamer-assisted CRISPR/Cas9 systems represent another technological shift, where engineered RNA aptamers function as transcriptional reporters without affecting Cas9 activity [74]. These systems enable precise genome editing and mutant screening with minimal off-target effects, offering new paradigms for functional analysis of gene family members without amplification-related artifacts.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for Mg²⁺ Optimization in Multi-Gene Family Studies
| Reagent/Equipment | Function | Application Notes |
|---|---|---|
| MgCl₂ stock solution (25 mM) | PCR cofactor source | Prepare nuclease-free, filter-sterilized aliquots |
| Mg²⁺-free PCR buffer | Reaction baseline | Enables precise Mg²⁺ titration |
| Proofreading polymerases (Pfu, Phusion) | High-fidelity amplification | Essential for accurate diversity assessment |
| Betaine (5M stock) | GC-rich template enhancer | Improves amplification of complex gene families |
| dNTP mix (10mM each) | Reaction substrates | Maintain constant concentration during Mg²⁺ optimization |
| Gradient thermocycler | Temperature optimization | Enables parallel testing of annealing stringency |
| Qubit fluorometer | DNA quantification | Accurate template normalization |
This systematic comparison demonstrates that Mg²⁺ concentration significantly influences PCR specificity, fidelity, and detection capabilities in multi-gene family analysis. The experimental data reveal that most applications achieve optimal performance between 1.5-2.5 mM Mg²⁺, with proofreading polymerases providing superior fidelity for diversity assessment. Researchers should employ empirical optimization using the provided protocols when characterizing new gene families, as sequence composition and primer characteristics create system-specific requirements. The continued development of engineered enzymes and amplification-free detection platforms promises to further mitigate Mg²⁺-dependent artifacts, enhancing the accuracy of gene family diversity studies and expanding our understanding of complex genomes.
In polymerase chain reaction (PCR) experiments, the accuracy with which a DNA polymerase replicates a template sequence—a property known as fidelity—is paramount for the success of downstream applications. The choice between standard Taq and high-fidelity proofreading enzymes carries significant implications for data integrity, particularly in sensitive contexts like cloning and sequencing. Furthermore, reaction conditions, specifically magnesium ion (Mg²⁺) concentration, are not merely a requirement for polymerase activity but a critical factor modulating enzymatic accuracy. This guide objectively compares the fidelity of standard Taq and high-fidelity polymerases, situating the analysis within broader research on how magnesium concentrations influence PCR fidelity. We present experimental data and methodologies to equip researchers and drug development professionals with the evidence needed to select and optimize enzymes for their specific applications.
Defining DNA Polymerase Fidelity
Polymerase fidelity refers to the accuracy of DNA sequence replication, measured as the error rate of misincorporated nucleotides per base per doubling event [75]. This accuracy is critical for the faithful transfer of genetic information and is expressed either as an absolute value (e.g., errors per thousand nucleotides) or relative to Taq DNA polymerase (1X) [75]. DNA polymerase fidelity is governed by two primary mechanisms: intrinsic nucleotide selectivity and proofreading activity.
- Intrinsic Nucleotide Selectivity: All DNA polymerases possess an active site that preferentially binds and incorporates correctly paired nucleotides. The geometry of this site ensures optimal catalytic efficiency only when Watson-Crick base pairing occurs. An incorrect nucleotide leads to a sub-optimal architecture, slowing incorporation and increasing the chance for the incorrect nucleotide to dissociate before the polymerase proceeds [75] [76].
- Proofreading Activity (3´→5´ Exonuclease): High-fidelity polymerases contain a separate 3´→5´ exonuclease domain that confers proofreading capability. When a mispaired nucleotide is incorporated, it causes a structural perturbation. The polymerase detects this, pauses synthesis, and moves the growing DNA strand into the exonuclease domain, where the incorrect base is excised. The correct nucleotide is then inserted, dramatically reducing the error rate [75] [19].
Comparative Fidelity Analysis: Taq vs. High-Fidelity Enzymes
The fidelity difference between standard non-proofreading polymerases like Taq and high-fidelity proofreading enzymes is substantial, spanning several orders of magnitude. The table below summarizes key performance metrics and characteristics.
Table 1: Comprehensive Comparison of Standard Taq and High-Fidelity DNA Polymerases
| Characteristic | Standard Taq Polymerase | High-Fidelity Polymerases (e.g., Q5, Pfu) |
|---|---|---|
| Proofreading Activity | No (lacks 3'→5' exonuclease) [77] | Yes (contains 3'→5' exonuclease) [19] [77] |
| Error Rate (per base per doubling) | ~1.5 × 10⁻⁴ [75] | Q5: ~5.3 × 10⁻⁷; Pfu: ~5.1 × 10⁻⁶ [75] |
| Relative Fidelity (vs. Taq) | 1X | Q5: 280X; Pfu: 30X [75] |
| Accuracy (1 base error per X bases) | ~6,500 bases [75] | Q5: ~1.87 million bases; Pfu: ~195,000 bases [75] |
| Primary Error Type | Base substitutions [75] | Base substitutions, with fewer overall errors [75] |
| Typical Extension Rate | ~150 nucleotides/second [77] | ~25 nucleotides/second [77] |
| "A-Tailing" Activity | Yes, produces PCR products with 3'A-overhangs [77] | No/Weak, typically produces blunt ends [77] |
| Ideal Application | Routine PCR, genotyping, gel analysis | Cloning, sequencing, site-directed mutagenesis, NGS library prep [76] [19] |
Quantitative data from PacBio SMRT sequencing, a method with a low background error rate suitable for measuring high-fidelity enzymes, reveals the stark contrast in performance [75]. While Taq polymerase has an error rate of approximately 1 in 6,500 bases, proofreading enzymes like Q5 and Pfu are far more accurate. The presence of a proofreading domain provides a significant advantage; for example, the exonuclease-deficient Deep Vent (exo-) polymerase has an error rate of 5.0 × 10⁻⁴, but the exonuclease-proficient version shows a 125-fold improvement, with an error rate of 4.0 × 10⁻⁶ [75].
Methodologies for Measuring Fidelity
Accurately quantifying polymerase error rates requires robust experimental assays. The evolution of these methods has increased the statistical power and accuracy of fidelity measurements.
Table 2: Key Experimental Methods for Assessing DNA Polymerase Fidelity
| Method | Principle | Workflow | Advantages & Limitations |
|---|---|---|---|
| Blue/White Colony Screening (lacZα) | PCR-amplified lacZα gene is cloned. Errors inactivate β-galactosidase, resulting in white colonies instead of blue [75] [19]. | 1. Amplify lacZα with test polymerase.2. Clone product into vector.3. Transform bacteria.4. Count blue vs. white colonies. | Advantage: High-throughput, cost-effective.Limitation: Only detects mutations that disrupt LacZ function; cannot identify all error types or locations [75]. |
| Sanger Sequencing | Individual cloned PCR products are directly sequenced and compared to the known template sequence [75]. | 1. Amplify target with test polymerase.2. Clone products.3. Pick individual colonies and Sanger sequence.4. Align sequences to reference. | Advantage: Detects all mutation types within sequenced reads.Limitation: Lower throughput and higher cost than colony screening; limits the total number of nucleotides sequenced [75]. |
| Next-Generation Sequencing (NGS) | PCR products are directly sequenced using high-throughput platforms (e.g., Illumina, PacBio), generating millions of reads [75] [25]. | 1. Amplify target with test polymerase.2. Prepare NGS library (may include molecular barcoding).3. Perform high-throughput sequencing.4. Map reads and call variants against reference. | Advantage: Extremely high throughput; provides a comprehensive mutation spectrum and rate with high statistical confidence.Limitation: Complex data analysis; potential for sequencing-derived errors requires controlled bioinformatics [75]. |
The following diagram illustrates a generalized workflow for a high-fidelity measurement experiment using modern sequencing technologies.
The Influence of Magnesium Concentration on Fidelity
Magnesium is an essential cofactor for DNA polymerase activity, but its concentration directly impacts fidelity. While standard PCR protocols often use optimized concentrations of 1.5-2.0 mM, these levels can be higher than physiological free Mg²⁺ levels (~0.5 mM) [78] [25].
Research on reverse transcriptases provides a clear model for how Mg²⁺ affects fidelity. Studies show that HIV-1 reverse transcriptase exhibits higher fidelity at physiological Mg²⁺ concentrations (0.5 mM) compared to standard reaction conditions (6 mM). In one experiment, a Next-Generation Sequencing assay revealed an approximately four-fold increase in mutations when Mg²⁺ was elevated from 0.5 mM to 6 mM [25]. This effect is not universal for all polymerases; for example, Moloney murine leukaemia virus (MuLV) and avian myeloblastosis virus (AMV) RTs showed unchanged fidelity across different Mg²⁺ concentrations [25]. This indicates that the impact of Mg²⁺ is enzyme-specific.
The mechanism is thought to involve Mg²⁺'s role in catalysis. Higher concentrations may relax the enzyme's stringency, increasing the rate of misincorporation by stabilizing the binding of incorrect nucleotides. Therefore, optimizing Mg²⁺ concentration is a critical step in reaction setup, especially for applications requiring utmost accuracy [78] [25].
The Scientist's Toolkit: Essential Reagents for Fidelity Research
The following table details key reagents and materials required for conducting polymerase fidelity experiments.
Table 3: Research Reagent Solutions for Fidelity Experiments
| Reagent/Material | Function in Experiment | Example Use Case |
|---|---|---|
| High-Fidelity DNA Polymerase | The enzyme under test, possessing proofreading activity for high-accuracy amplification. | Q5 High-Fidelity DNA Polymerase for generating amplicons with ultra-low error rates for sequencing [75]. |
| Standard Taq Polymerase | Reference enzyme for establishing a baseline fidelity level (1X). | Used as a control in fidelity assays to calculate relative improvement of high-fidelity enzymes [75]. |
| Control Plasmid (e.g., lacZα) | A well-characterized DNA template with a known sequence for error detection. | pBS▽EcoRV567 plasmid used as a template in lacZα-based α-complementation assays [25]. |
| MgCl₂ Solution | A variable cofactor to study its specific effect on polymerase error rate. | Used in concentration gradients (e.g., 0.5 mM to 6 mM) to assess fidelity under different conditions [25]. |
| NGS Library Prep Kit | Prepares PCR amplicons for high-throughput sequencing by fragmenting, sizing, and adding platform-specific adapters. | NEBNext Ultra II DNA Library Prep Kit for Illumina used for preparing fidelity assay products for sequencing [25]. |
| PCR Additives (e.g., DMSO) | Additives that help denature complex templates, improving amplification efficiency and yield. | 5% DMSO used to successfully amplify an extremely GC-rich EGFR promoter sequence [29]. |
The selection between standard Taq and high-fidelity DNA polymerases is a critical decision that directly impacts the validity of experimental results. High-fidelity proofreading enzymes, with error rates up to 280 times lower than Taq, are indispensable for applications where sequence integrity is paramount. Furthermore, this comparison underscores that fidelity is not an immutable property of the enzyme alone. Reaction conditions, particularly the concentration of Mg²⁺, serve as a powerful modulator of polymerase accuracy. The evidence shows that for some enzymes, moving from standardized, high Mg²⁺ concentrations to more physiological levels can significantly enhance replication fidelity. For researchers in drug development and basic science, a rigorous approach that pairs the appropriate high-fidelity enzyme with optimized, physiologically relevant reaction conditions will yield the most reliable and reproducible genetic data.
In the realm of molecular biology, the pursuit of reproducible, high-fidelity amplification represents a cornerstone of reliable research and diagnostic applications. While numerous factors influence polymerase chain reaction (PCR) outcomes, magnesium chloride (MgCl₂) concentration emerges as perhaps the most pivotal variable governing both reaction efficiency and amplification accuracy. As a cofactor for thermostable DNA polymerases, Mg²⁺ facilitates enzyme activity and primer-template interactions, yet its concentration must be precisely calibrated to avoid detrimental effects on fidelity. The establishment of validated Mg²⁺ ranges is thus not merely a technical consideration but a fundamental prerequisite for scientific rigor in molecular applications ranging from clinical diagnostics to large-scale cloning projects.
The critical importance of Mg²⁺ optimization extends beyond basic amplification efficiency to encompass the very accuracy of DNA replication. Recent investigations have revealed that Mg²⁺ concentrations significantly influence polymerase error rates, with profound implications for applications requiring high precision, such as mutant detection, sequencing, and cloning. This guide synthesizes current experimental evidence to establish validated Mg²⁺ parameters that support reproducible, high-fidelity amplification across diverse PCR applications and template types.
Magnesium-Dependent Fidelity Mechanisms: Biochemical Foundations
The fidelity of DNA polymerase enzymes is intrinsically linked to Mg²⁺ concentration through well-defined biochemical mechanisms. Magnesium ions participate directly in the catalytic center of DNA polymerases, facilitating the nucleotidyl transfer reaction by coordinating the interaction between the enzyme, template DNA, primer terminus, and incoming dNTPs. This coordination involves two metal ions (typically Mg²⁺) that position the reactive groups for catalysis and stabilize the transition state.
Beyond its catalytic role, Mg²⁺ influences reaction fidelity through modulation of enzyme processivity, DNA melting characteristics, and primer-template stability. Elevated Mg²⁺ concentrations have been demonstrated to reduce the base substitution fidelity of certain DNA polymerases, including HIV-1 reverse transcriptase, by affecting the enzyme's ability to discriminate against incorrect nucleotides [25]. This phenomenon underscores the importance of optimizing Mg²⁺ concentrations to balance amplification efficiency with accuracy requirements.
Table 1: Magnesium-Dependent Effects on PCR Components
| PCR Component | Low Mg²⁺ Effect | High Mg²⁺ Effect |
|---|---|---|
| DNA Polymerase Activity | Reduced catalytic efficiency | Increased activity, potentially reduced fidelity |
| Primer-Template Stability | Decreased annealing efficiency | Increased non-specific binding |
| DNA Melting Temperature | Lower Tm requiring optimization | Increased Tm affecting denaturation |
| Reaction Fidelity | Potentially higher fidelity for some enzymes | Possibly reduced fidelity with increased errors |
| Product Specificity | Increased stringency | Decreased stringency with spurious products |
The relationship between Mg²⁺ concentration and amplification fidelity varies among polymerase families. Proofreading enzymes with 3'→5' exonuclease activity, such as Pfu and Pwo polymerases, generally maintain higher fidelity across a broader Mg²⁺ range compared to non-proofreading enzymes like Taq polymerase [15]. However, even high-fidelity enzymes exhibit concentration-dependent error rates, necessitating empirical optimization for critical applications.
Quantitative Mg²⁺ Optimization: Evidence-Based Concentration Ranges
Comprehensive Meta-Analysis of Mg²⁺ Concentrations
A systematic meta-analysis of 61 peer-reviewed studies published between 1973 and 2024 provides robust, quantitative guidance for Mg²⁺ optimization in PCR [5]. This comprehensive analysis revealed a strong logarithmic relationship between MgCl₂ concentration and DNA melting temperature, establishing an evidence-based optimal range of 1.5 to 3.0 mM for standard amplification protocols. Within this range, every 0.5 mM increase in MgCl₂ concentration was associated with a 1.2°C increase in melting temperature, providing a predictable framework for protocol adjustment.
The meta-analysis further demonstrated that template characteristics significantly influence optimal Mg²⁺ requirements. Specifically, genomic DNA templates consistently required higher Mg²⁺ concentrations than simpler plasmid templates, reflecting the differential Mg²⁺ demands of complex versus straightforward templates. This template-specific effect underscores the limitation of one-size-fits-all Mg²⁺ formulations and emphasizes the need for application-specific optimization.
Polymerase-Specific Magnesium Optimization
Different DNA polymerases exhibit distinct Mg²⁺ optima and fidelity profiles, necessitating enzyme-specific optimization strategies. Experimental comparisons of error rates across six commonly used DNA polymerases revealed substantial variation in fidelity, with high-fidelity enzymes such as Pfu, Phusion, and Pwo demonstrating error rates more than 10-fold lower than Taq polymerase [15].
Table 2: Polymerase-Specific Error Rates and Magnesium Considerations
| DNA Polymerase | Error Rate (errors/bp/duplication) | Fidelity Relative to Taq | Mg²⁺ Considerations |
|---|---|---|---|
| Taq | 1–20 × 10⁻⁵ | 1x | Standard optimization (1.5-2.5 mM) |
| AccuPrime-Taq HF | ~1.0 × 10⁻⁵ | 9x better | Follow manufacturer recommendations |
| KOD Hot Start | ~10⁻⁶ range | 50x better | Often requires higher Mg²⁺ (2.0-3.0 mM) |
| Pfu | 1-2 × 10⁻⁶ | 6-10x better | Broader Mg²⁺ tolerance (1.5-3.0 mM) |
| Phusion Hot Start | 4 × 10⁻⁷ (HF buffer) | >50x better | Buffer-dependent optimization |
| Pwo | ~10⁻⁶ range | >10x better | Similar to Pfu requirements |
The experimental data demonstrate that while high-fidelity polymerases generally maintain superior accuracy across Mg²⁺ concentrations, optimal fidelity is achieved within manufacturer-recommended ranges that typically fall between 1.5 and 2.5 mM for standard amplification [15]. Deviation from these ranges, particularly excess Mg²⁺, can diminish the fidelity advantage of proofreading enzymes.
Experimental Protocols for Magnesium Optimization
Systematic Mg²⁺ Titration Methodology
A standardized approach to Mg²⁺ optimization ensures reproducible, high-fidelity amplification across diverse applications. The following protocol, synthesized from multiple experimental studies, provides a robust framework for establishing laboratory-specific Mg²⁺ parameters:
Reaction Setup:
- Prepare a master mix containing all reaction components except MgCl₂ and template
- Aliquot equal volumes into individual reaction tubes
- Add MgCl₂ from a stock solution to create a concentration series spanning 0.5 mM to 5.0 mM in 0.5 mM increments
- Include negative controls (no template) for each Mg²⁺ concentration
- Use a standardized template of known concentration and complexity
Thermocycling Parameters:
- Employ a touchdown or gradient protocol to account for Mg²⁺-dependent Tm effects
- Include sufficient extension time for complete product synthesis
- Limit cycle number to 25-30 to minimize accumulation of stochastic errors
Analysis and Interpretation:
- Resolve amplification products by agarose gel electrophoresis
- Quantify product yield using fluorescent DNA-binding dyes
- Assess specificity by band intensity and absence of spurious products
- Select the lowest Mg²⁺ concentration that provides robust, specific amplification
This methodological framework aligns with optimization approaches validated in multiple experimental contexts [79] [5]. For applications requiring maximal fidelity, such as cloning or sequencing, additional validation through replicate reactions and downstream sequence analysis is recommended.
Fidelity Assessment Techniques
Beyond optimization for efficiency, specific methodologies enable quantitative assessment of PCR fidelity under different Mg²⁺ conditions. The lacZα-based α-complementation assay represents a robust approach for fidelity determination, measuring mutation rates through functional selection [15] [25]. This method employs a two-round DNA synthesis approach using an RNA template, with mutation rates calculated based on the frequency of loss-of-function clones.
Next Generation Sequencing (NGS) approaches provide complementary fidelity assessment, enabling comprehensive mutation profiling across entire amplicons. Experimental applications of NGS fidelity analysis have demonstrated approximately four-fold increases in mutation rates for HIV-1 reverse transcriptase under high Mg²⁺ (6 mM) conditions compared to physiological Mg²⁺ (0.5 mM) concentrations [25]. This methodology offers unprecedented resolution for characterizing Mg²⁺-dependent error profiles.
Comparative Performance Across PCR Applications
Application-Specific Magnesium Optimization
The optimal Mg²⁺ concentration varies significantly across PCR applications, reflecting differences in template characteristics, enzyme selection, and detection requirements. Experimental evidence supports the following application-specific guidelines:
Standard PCR:
- Mg²⁺ range: 1.5-2.5 mM
- Priority: Robustness and general amplification efficiency
- Considerations: Template complexity dictates specific requirements, with genomic DNA typically requiring higher concentrations [79] [5]
Quantitative PCR (qPCR):
- Mg²⁺ range: 2.0-3.0 mM
- Priority: Sensitivity and fluorescent detection compatibility
- Considerations: Increased Mg²⁺ may enhance signal intensity but requires careful optimization to maintain specificity [79]
Reverse Transcription PCR (RT-PCR):
- Mg²⁺ range: 2.5-3.5 mM
- Priority: High cDNA yield and minimal RNase activity
- Considerations: Combined reverse transcription and amplification steps may require elevated Mg²⁺ [79]
High-Throughput Cloning:
- Mg²⁺ range: 1.5-2.0 mM
- Priority: Maximal fidelity with high-fidelity polymerases
- Considerations: Lower Mg²⁺ may enhance fidelity for some enzyme systems [15] [25]
Template-Dependent Magnesium Requirements
Template characteristics significantly influence optimal Mg²⁺ concentrations, with complex templates generally requiring higher concentrations than simple templates. The meta-analysis by [5] quantitatively established that genomic DNA templates consistently require higher Mg²⁺ concentrations than plasmid templates. This template effect reflects the differential impact of Mg²⁺ on DNA melting and enzyme processivity in complex versus simple sequences.
GC-rich templates present particular optimization challenges, often benefiting from elevated Mg²⁺ concentrations (2.5-3.5 mM) or specialized buffer formulations. Experimental evidence indicates that Mg²⁺ concentrations at the higher end of the optimal range stabilize GC-rich duplexes, improving amplification efficiency without necessarily compromising fidelity when using high-fidelity enzymes [79] [5].
The Scientist's Toolkit: Essential Reagents for Fidelity-Optimized PCR
Table 3: Research Reagent Solutions for Magnesium-Optimized PCR
| Reagent Category | Specific Examples | Function in Fidelity Optimization |
|---|---|---|
| High-Fidelity DNA Polymerases | Pfu, Phusion, KOD | Proofreading activity reduces errors; defined Mg²⁺ optima |
| Magnesium Chloride Solutions | Molecular grade MgCl₂ | Precise concentration adjustment; lot-to-lot consistency |
| Optimization Buffers | Mg²⁺-free buffers with additives | Systematic titration without component interference |
| dNTP Mixtures | Purified, balanced dNTPs | Prevent Mg²⁺ sequestration; ensure equitable incorporation |
| Template Preparation Kits | Column-based purification | Remove contaminants that alter Mg²⁺ availability |
| Fidelity Assessment Systems | lacZα complementation assays | Quantitative error rate measurement |
| Additive Solutions | DMSO, betaine, BSA | Modulate Mg²⁺ effects on specific templates |
The selection of appropriate reagent systems fundamentally influences the success of Mg²⁺ optimization efforts. High-fidelity DNA polymerases with proofreading activity (3'→5' exonuclease) provide inherent error correction mechanisms that reduce mutation frequencies, particularly when optimized with appropriate Mg²⁺ concentrations [15]. Purified nucleotide preparations prevent Mg²⁺ sequestration by contaminating pyrophosphates, while specialized buffer systems enable precise Mg²⁺ titration without confounding variables.
Visualization of Magnesium Optimization Workflows
Experimental Optimization Pathway
Magnesium's Mechanism in PCR Fidelity
The establishment of validated Mg²⁺ concentration ranges represents a critical advancement in the pursuit of reproducible, high-fidelity PCR amplification. Experimental evidence consistently supports an optimal Mg²⁺ range of 1.5-3.0 mM, with specific concentrations determined by template characteristics, polymerase selection, and application requirements. The demonstrated influence of Mg²⁺ on polymerase error rates underscores the importance of systematic optimization beyond mere amplification efficiency.
Future directions in Mg²⁺ optimization will likely incorporate polymerase-specific fidelity profiles under physiological Mg²⁺ conditions, advanced computational modeling of Mg²⁺-dependent amplification kinetics, and specialized formulations for emerging applications such as long-amplicon PCR and multiplexed detection systems. By implementing the evidence-based guidelines presented herein, researchers can achieve significant improvements in amplification reliability, data quality, and experimental reproducibility across diverse molecular applications.
Conclusion
The concentration of magnesium chloride is a pivotal, yet often empirically adjusted, parameter that directly governs the success, specificity, and fidelity of PCR. A thorough understanding of its foundational role as a polymerase cofactor, combined with a systematic methodological approach to optimization, allows researchers to effectively troubleshoot common amplification issues. The validation data consistently demonstrates that moving away from a one-size-fits-all concentration towards a finely tuned, template- and application-specific Mg²⁺ level is crucial for achieving high fidelity. For the biomedical and clinical research community, adopting these evidence-based optimization strategies is imperative for enhancing the reliability of downstream applications—from accurate genotyping and variant detection in diagnostics to ensuring the integrity of clones in therapeutic development. Future work should focus on integrating computational and AI-driven models to predict optimal Mg²⁺ conditions, further bridging the gap between in vitro efficiency and physiological relevance.
References
- 1. What Is the Role of MgCl2 in PCR Amplification Reactions? [https://www.excedr.com/resources/what-is-the-role-of-mgcl2-in-pcr]
- 2. PCR Setup—Six Critical Components to Consider [https://www.thermofisher.com/us/en/home/life-science/cloning/cloning-learning-center/invitrogen-school-of-molecular-biology/pcr-education/pcr-reagents-enzymes/pcr-component-considerations.html]
- 3. Distinct Roles of the Active-site Mg2+ Ligands, Asp882 and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3030377/]
- 4. Role of magnesium ions in the reaction mechanism at the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4939058/]
- 5. Investigating concentration effects on reaction efficiency ... [https://pubmed.ncbi.nlm.nih.gov/40436087/]
- 6. How does magnesium concentration affect PCR? [https://www.aatbio.com/resources/faq-frequently-asked-questions/how-does-magnesium-concentration-affect-pcr]
- 7. Smearing in PCR Amplification | NTA NET LIFE SCIENCE [https://www.letstalkacademy.com/smearing-in-pcr-amplification-due-to-low-mg-ions-concentration/]
- 8. What is Magnesium Chloride and Why Is It Important for ... [https://www.linkedin.com/posts/dip-ghosh-2329b3291_pcr-mgcl2-molecularbiology-activity-7356124849922035713-5KhE]
- 9. PCR conditions | Primer annealing specificity | PCR buffers [https://www.qiagen.com/us/knowledge-and-support/knowledge-hub/bench-guide/pcr/introduction/pcr-conditions]
- 10. Kinetic and thermodynamic analysis defines roles for two ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7948414/]
- 11. Stepwise kinetic equilibrium models of quantitative ... [https://bmcbioinformatics.biomedcentral.com/articles/10.1186/1471-2105-13-203]
- 12. Engineering of novel DNA polymerase variants for single ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12274487/]
- 13. Recent advances in CRISPR-based single-nucleotide fidelity ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12219407/]
- 14. What does DNA polymerase fidelity mean? [https://www.neb.com/en-us/tools-and-resources/feature-articles/polymerase-fidelity-what-is-it-and-what-does-it-mean-for-your-pcr?srsltid=AfmBOor0tancQMSA0CDOLZczw_TMjfWJr7f4frzOaG1mcfz4N2SUAqAA]
- 15. Error Rate Comparison during Polymerase Chain Reaction ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4150459/]
- 16. The A, B, C, D's of replicative DNA polymerase fidelity [https://pmc.ncbi.nlm.nih.gov/articles/PMC12634111/]
- 17. Human Immunodeficiency Virus Reverse Transcriptase ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4135932/]
- 18. PCR Optimization: Factors Affecting Reaction Specificity ... [https://www.mybiosource.com/learn/pcr-optimization-factors-affecting-reaction-specificity-and-efficien/]
- 19. DNA Polymerase—Four Key Characteristics for PCR [https://www.thermofisher.com/us/en/home/life-science/cloning/cloning-learning-center/invitrogen-school-of-molecular-biology/pcr-education/pcr-reagents-enzymes/dna-polymerase-characteristics.html]
- 20. Investigating concentration effects on reaction efficiency ... [https://www.sciencedirect.com/science/article/abs/pii/S0003269725001472]
- 21. Development and comparative evaluation of LAMP, nested ... [https://bmcmicrobiol.biomedcentral.com/articles/10.1186/s12866-025-04295-8]
- 22. Optimization of loop-mediated isothermal amplification ... [https://www.nature.com/articles/s41598-024-72228-y]
- 23. Engineering of novel DNA polymerase variants for single ... [https://www.nature.com/articles/s41598-025-10211-x]
- 24. PCR Quality Control: Comprehensive Guide to Accuracy ... [https://goldengatebio.com/pcr-quality-control-comprehensive-guide-to-accuracy-validation-and-reproducibility-in-molecular-biology/]
- 25. Physiological magnesium concentrations increase fidelity ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10019084/]
- 26. Mg2+ dependency of HIV-1 reverse transcription, inhibition ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC1325016/]
- 27. Alternative divalent cations (Zn2+, Co2+, and Mn2+) are not ... [https://bmcbiochem.biomedcentral.com/articles/10.1186/s12858-015-0041-x]
- 28. Guidelines for PCR Optimization with Taq DNA Polymerase [https://www.neb.com/en-us/tools-and-resources/usage-guidelines/guidelines-for-pcr-optimization-with-taq-dna-polymerase?srsltid=AfmBOorhsN2AhLiuqs-JndhLrRob-gAqjosZiFdwahp6qBUPTPX8le47]
- 29. Optimization of PCR Conditions for Amplification of GC‐ ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6807403/]
- 30. Polymerase Chain Reaction: Basic Protocol Plus ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4846334/]
- 31. Four tips for PCR amplification of GC-rich sequences [https://www.neb.com/en-us/nebinspired-blog/four-tips-for-pcr-amplification-of-gc-rich-sequences?srsltid=AfmBOoqzPZRflJ4jssWPn41_qJFKZ6YG_p0_8v-r6MUa5SvmplA_9L-C]
- 32. StellarTaq DNA Polymerase [https://www.watchmakergenomics.com/enzymes-and-proteins/molecular-diagnostics/stellartaq-dna-polymerase.html]
- 33. MIQE 2.0: Revision of the Minimum Information for ... [https://pubmed.ncbi.nlm.nih.gov/40272429/]
- 34. MIQE Guidelines | Thermo Fisher Scientific - US [https://www.thermofisher.com/us/en/home/life-science/pcr/real-time-pcr/real-time-pcr-assays/why-choose-taqman-assays/publish-real-time-pcr-results.html]
- 35. Optimization of the real-time PCR platform method using ... [https://www.nature.com/articles/s41598-025-16972-9]
- 36. Establishment of a simple prediction method for DNA melting ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0321885]
- 37. What is the role of magnesium in PCR, and what ... [https://help.takarabio.com/36168/kb/article/110034/what-is-the-role-of-magnesium-in-pcr-and-what-is-the-optimal-concentration]
- 38. Mathematical Modeling and Thermodynamic Integration for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12620865/]
- 39. Four tips for PCR amplification of GC-rich sequences [https://www.neb.com/en-us/nebinspired-blog/four-tips-for-pcr-amplification-of-gc-rich-sequences?srsltid=AfmBOop5xopx4tFOq4R-bt8qYdOkzP4d4sp_6D5mxoe_wO5jPf4T3ZdJ]
- 40. How to Sequence a Gene: Step-by-Step Experimental ... [https://www.cd-genomics.com/resource-how-to-sequence-a-gene.html]
- 41. Determination of the stability constants of Mn2+ and Mg2+ ... [https://pubmed.ncbi.nlm.nih.gov/6254771/]
- 42. Guidelines for PCR Optimization with Taq DNA Polymerase [https://www.neb.com/en-us/tools-and-resources/usage-guidelines/guidelines-for-pcr-optimization-with-taq-dna-polymerase?srsltid=AfmBOoqWdL2x11z32juyrN6hoF8GTYg2ZJkNlxPT0pGf0niirFRCsChY]
- 43. Comparison of Magnesium and Manganese Ions on the ... [https://chemrxiv.org/engage/chemrxiv/article-details/67cf60e8fa469535b9ca6bbf]
- 44. Understanding the Buffer Chemistry in PCR Kit V2 [https://thearkdb.org/understanding-the-buffer-chemistry-in-pcr-kit-v2-how-it-improves-amplification-of-difficult-templates/]
- 45. EVO HS Super-Fidelity DNA Polymerase: The Ultimate High ... [https://www.umespe.com/evo-hs-super-fidelity-dna-polymerase-the-ultimate-high-fidelity-enzyme-for-precise-pcr/]
- 46. Variants of PCR [https://grokipedia.com/page/Variants_of_PCR]
- 47. Optimizing PCR Reaction Conditions for High Fidelity and ... [https://www.labx.com/resources/optimizing-pcr-reaction-conditions-for-high-fidelity-and-yield/5579]
- 48. Just What Do All These Additives Do? [https://bitesizebio.com/19420/just-what-do-all-these-additives-do/]
- 49. Four tips for PCR amplification of GC-rich sequences [https://www.neb.com/en-us/nebinspired-blog/four-tips-for-pcr-amplification-of-gc-rich-sequences?srsltid=AfmBOooPiYR4t8AE97nGrQfEpMLYRkhtAwB0SIcNLY1Eb1GSxZW4bld8]
- 50. Impact of metal ions on PCR inhibition and RT- ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7782418/]
- 51. Number of PCR Cycles and Magnesium Chloride ... [https://file.scirp.org/Html/6-2270410_49726.htm]
- 52. Guidelines for PCR Optimization with Taq DNA Polymerase [https://www.neb.com/en-us/tools-and-resources/usage-guidelines/guidelines-for-pcr-optimization-with-taq-dna-polymerase?srsltid=AfmBOorhJe2lt5Etyx8wNCzk_E-dEXbKR-SfyGpJJjhQUaABedlYAs2c]
- 53. 2025 Oligonucleotide Design Guide: Complete User Manual [https://oligopool.com/resources/user-guide]
- 54. PCR inhibition in qPCR, dPCR and MPS—mechanisms and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7072044/]
- 55. Overcoming qPCR Inhibitors: Strategies for Reliable ... [https://www.promegaconnections.com/overcoming-qpcr-inhibitors-strategies-for-reliable-quantification/]
- 56. Evaluation of PCR-enhancing approaches to reduce ... [https://www.sciencedirect.com/science/article/abs/pii/S0048969724069250]
- 57. PCR Product Purification Techniques: Complete Guide [https://microzone.co.uk/complete-guide-pcr-product-purification-techniques/]
- 58. PCR Cleanup: Purifying Your PCR Product [https://www.thermofisher.com/us/en/home/references/an-introduction-to-cloning-a-researchers-guide-to-cloning-dna/purifying-your-pcr-product.html]
- 59. PCR optimization for buccal swab-derived samples [https://pubmed.ncbi.nlm.nih.gov/41272443/]
- 60. Essential Tips for PCR & DNA Cleanup: Maximize Your ... [https://www.neb.com/en-us/nebinspired-blog/essential-tips-for-pcr-and-dna-cleanup-maximize-your-dna-purity-and-yield?srsltid=AfmBOop3y0xM0o1u6ympBS-zodpJbbuKjNXBrJhXlDugTZ6rLsyicbSa]
- 61. 聚合酶链反应:基本协议 加 上故障排除和优化策略 [https://www.jove.com/cn/t/3998/polymerase-chain-reaction-basic-protocol-plus-troubleshooting]
- 62. DMSO and Betaine Greatly Improve Amplification of GC-Rich ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0011024]
- 63. CN101392282A - 一种扩增高gc含量片段的 pcr ... - Google Patents [https://patents.google.com/patent/CN101392282A/zh]
- 64. How to optimize PCR conditions for GC-rich regions? [https://synapse.patsnap.com/article/how-to-optimize-pcr-conditions-for-gc-rich-regions]
- 65. DMSO and betaine significantly enhance the PCR ... [https://pubmed.ncbi.nlm.nih.gov/32433893/]
- 66. What does DNA polymerase fidelity mean? [https://www.neb.com/en-us/tools-and-resources/feature-articles/polymerase-fidelity-what-is-it-and-what-does-it-mean-for-your-pcr?srsltid=AfmBOook4Su8P-jwPBw2wqbOZ829A4VMJq3eZiRVqCdD-zV6aFAorGmH]
- 67. Working with PCR [https://www.sigmaaldrich.com/US/en/technical-documents/technical-article/genomics/pcr/working-with-pcr?srsltid=AfmBOopCZNOYPmbcdXkNdUesnmm3IuyDsI5g-SdEIHtniLunWRNKQ9_M]
- 68. Next-generation sequencing-based analysis of reverse transcriptase... [https://pubmed.ncbi.nlm.nih.gov/28778390/]
- 69. What does DNA polymerase fidelity mean? [https://www.neb.com/en-us/tools-and-resources/feature-articles/polymerase-fidelity-what-is-it-and-what-does-it-mean-for-your-pcr?srsltid=AfmBOoqhEtgGU4skDeXB41ExCs209BSyVtRxn6NUDoHBTkBNb5qfBkW9]
- 70. Magnesium Induced Assembly of a Complete DNA ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC1868394/]
- 71. Effect of concentration of MgCl2 on random-amplified DNA ... [https://pubmed.ncbi.nlm.nih.gov/8024785/]
- 72. PCR amplification introduces errors into mononucleotide ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC1187094/]
- 73. NAPTUNE: nucleic acids and protein biomarkers testing ... [https://www.nature.com/articles/s41467-025-56653-9]
- 74. Development of an RNA aptamer-assisted CRISPR/Cas9 ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12614593/]
- 75. What does DNA polymerase fidelity mean? [https://www.neb.com/en-us/tools-and-resources/feature-articles/polymerase-fidelity-what-is-it-and-what-does-it-mean-for-your-pcr?srsltid=AfmBOor9GgjRZA2Uo51g0lq7kwRokS65EFu9YinonKIxh3Y7DGNf-C61]
- 76. High-fidelity DNA Polymerases & When to use them [https://clentlifescience.co.uk/high-fidelity-dna-polymerases-and-when-to-use-them/]
- 77. DNA polymerases used in PCR [https://www.qiagen.com/us/knowledge-and-support/knowledge-hub/bench-guide/pcr/introduction/enzymes-used-in-pcr]
- 78. Optimizing your PCR [https://www.takarabio.com/learning-centers/pcr/faq/optimization?srsltid=AfmBOoqFUektbIQC2CDFnmkfoOJEALZrQ7q9k8TAUEbKcpmR7sHu-CZT]
- 79. PCR Reagent Selection 101: Expert Tips for Accuracy and ... [https://prolabcorp.com/blog/pcr-reagent-selection-101-expert-tips-for-accuracy-and-efficiency/?srsltid=AfmBOoohGaEkjpt6j-4qhXViTtM3hwo3a2aCx5eqabtVDQ0Gp_eCxby9]