Dynamic Targeting: How STAT SH2 Domain Flexibility is Reshaping Drug Discovery
This article provides a comprehensive exploration of the molecular dynamics and conformational flexibility of STAT SH2 domains, a critical target in oncology and immunology.
Dynamic Targeting: How STAT SH2 Domain Flexibility is Reshaping Drug Discovery
Abstract
This article provides a comprehensive exploration of the molecular dynamics and conformational flexibility of STAT SH2 domains, a critical target in oncology and immunology. Aimed at researchers and drug development professionals, we first establish the foundational structural principles and unique characteristics of STAT-type SH2 domains. The piece then delves into advanced computational methodologies, including molecular dynamics simulations and virtual screening, that leverage this flexibility for inhibitor design. We address key challenges in simulating these dynamic systems and present optimization strategies to enhance predictive accuracy. Finally, the article covers rigorous validation frameworks, comparing STAT SH2 dynamics with other domains and evaluating emerging allosteric targeting approaches. By synthesizing foundational knowledge with cutting-edge applications, this review serves as a strategic guide for developing novel therapeutics that target the dynamic landscape of STAT signaling.
The Structural Blueprint and Innate Flexibility of STAT SH2 Domains
The Src Homology 2 (SH2) domain is a critical protein-protein interaction module found in numerous signaling proteins, including the Signal Transducer and Activator of Transcription 3 (STAT3) [1]. These domains function as fundamental "readers" of phosphotyrosine (pTyr) modifications, enabling the transduction of cellular signals that regulate processes such as proliferation, differentiation, and survival [1]. Among the SH2 domain-containing proteins, STAT3 has emerged as a particularly attractive therapeutic target in oncology due to its frequent constitutive activation in a wide range of human cancers, which is often associated with poor prognosis [2] [3] [4]. The canonical architecture of the SH2 domain, characterized by a conserved αβββα sandwich fold, contains specialized binding pockets that recognize phosphorylated tyrosine residues and their specific sequence contexts. This whitepaper provides an in-depth technical examination of this canonical architecture, with a specific focus on the pY+0 binding pocket of STAT3, and explores its implications for drug discovery within the broader context of molecular dynamics and SH2 domain flexibility research.
The Canonical SH2 Domain Fold
Structural Organization of the αβββα Sandwich
The SH2 domain adopts a conserved tertiary structure known as an αβββα sandwich or fold [5] [6]. This canonical architecture consists of a central anti-parallel β-sheet flanked by two α-helices, forming a scaffold that is both structurally stable and functionally versatile [5] [1]. As illustrated in Figure 1, the core fold comprises:
- A Central β-Sheet: Formed by three anti-parallel β-strands (typically labeled βB, βC, and βD) [5].
- Flanking α-Helices: Designated αA and αB, which pack against opposite faces of the β-sheet, creating the characteristic "sandwich" structure [5] [1].
- Stability and Conservation: This compact globular fold, comprising approximately 80-350 amino acids, provides a stable platform for the specific recognition of phosphotyrosine-containing motifs [7] [1].
Figure 1: The canonical αβββα sandwich fold of the SH2 domain.
STAT3 SH2 Domain Architecture
Within the STAT3 protein, the SH2 domain (residues 586-690) plays an indispensable role in both its recruitment to activated receptor complexes and its subsequent homodimerization [3] [4]. The STAT3 monomer, as extracted from the 1BG1 crystal structure, reveals multiple domains:
- N-terminal four-helix bundle (residues 138-320)
- Eight-stranded β-barrel (residues 321-465)
- α-helical linker domain (residues 466-585)
- SH2 domain (residues 586-690)
- Loop domain (residues 691-715) containing the critical Tyr705 phosphorylation site [3] [4]
Activation involves phosphorylation of Tyr705 within the loop domain, creating a phosphotyrosine motif that binds in trans to the SH2 domain of another STAT3 monomer, facilitating active dimer formation and subsequent nuclear translocation [3] [5] [4].
The pY+0 Binding Pocket: Structure and Function
Anatomical Organization of the Binding Groove
The phosphopeptide-binding groove of the STAT3 SH2 domain is strategically located on its surface and can be divided into distinct sub-pockets that recognize specific residues flanking the phosphotyrosine. These sub-pockets provide both binding affinity and sequence specificity [5].
Table 1: Key Binding Pockets in the STAT3 SH2 Domain
| Binding Pocket | Structural Role | Key Residues | Functional Significance |
|---|---|---|---|
| pY+0 | Binds phosphotyrosine (pTyr705) | R609, S613 | Critical for STAT3 dimerization; primary target for inhibitors |
| pY+1 | Binds hydrophobic residue at pTyr+1 position | L706 | Contributes to binding specificity and affinity |
| Hydrophobic Side | Accommodates hydrophobic residues | Various | Enhances binding stability and specificity |
The pY+0 pocket represents the primary binding site for the phosphorylated tyrosine (pTyr705) and is therefore absolutely essential for STAT3 activation through dimerization [5]. Key residues within this pocket, particularly Arg609 and Ser613, form critical interactions with the phosphate group of pTyr705, enabling high-affinity binding [2] [3] [4].
Molecular Recognition Mechanism
The molecular recognition within the pY+0 pocket involves specific, well-characterized interactions:
- Electrostatic Complementarity: The positively charged guanidinium group of Arg609 forms strong salt bridges with the negatively charged phosphate group of pTyr705 [3] [4].
- Hydrogen Bonding Networks: Ser613 and other polar residues form additional hydrogen bonds with the phosphate moiety, enhancing binding specificity and affinity [3] [5].
- Shape Complementarity: The pocket architecture provides a sterically constrained environment that preferentially accommodates phosphorylated tyrosine over other residues [5].
This sophisticated recognition mechanism ensures that STAT3 dimerization occurs specifically in response to proper activation signals, maintaining the fidelity of cellular signaling.
Molecular Dynamics and Domain Flexibility
Conformational Flexibility of the SH2 Domain
Traditional structure-based drug design approaches often rely on static crystal structures, which may not fully capture the dynamic behavior of proteins in solution. The STAT3 SH2 domain exhibits significant conformational flexibility, particularly in its phosphopeptide-binding region [3] [4]. Key observations include:
- In crystal structures (e.g., 1BG1), the phosphopeptide binding region is resolved to only approximately 20 Å due to inherent conformational flexibility [3] [4].
- Molecular dynamics (MD) simulations reveal that the SH2 domain undergoes substantial conformational fluctuations that influence ligand binding [3] [4].
- The domain exhibits "induced fit" binding characteristics, where ligand binding actively reshapes the binding pocket architecture [3].
Advanced Simulation Approaches
To address the challenges posed by SH2 domain flexibility, researchers have developed sophisticated simulation methodologies:
- Molecular Dynamics (MD) Simulations: Kong et al. conducted MD simulations of the STAT3 SH2 domain in complex with CJ-887 (a high-affinity peptidomimetic), observing ligand-induced conformational changes that enhance binding [2] [3] [4].
- Induced-Active Site Models: Averaged structures from MD trajectories can be used as "induced-active site" receptor models for more accurate virtual screening [3] [4].
- Enhanced Sampling Techniques: Advanced methods such as Gaussian-accelerated MD and metadynamics provide more comprehensive exploration of conformational landscapes [3] [5].
Figure 2: Molecular dynamics workflow for studying SH2 domain flexibility.
Experimental and Computational Methodologies
Structure-Based Virtual Ligand Screening
The integration of molecular dynamics with structure-based virtual ligand screening (SB-VLS) represents a powerful approach for identifying novel STAT3 inhibitors [3] [4]:
- Receptor Model Preparation: Using averaged MD structures rather than static crystal structures as receptor models for docking [3] [4].
- Library Screening: Virtual screening of compound libraries (e.g., SPECS database with ~110,000 compounds) using docking algorithms [2] [3].
- Hierarchical Refinement: Initial screening followed by re-docking and re-scoring of top hits (e.g., top 30%) with more precise algorithms [3] [4].
- Interaction-Based Selection: Prioritizing compounds that form specific interactions with key pY+0 pocket residues (Arg609, Ser613) [2] [3].
Natural Product Screening
Recent studies have explored natural product libraries for STAT3-SH2 domain inhibitors [5] [8]:
- Library Preparation: 182,455 natural compounds from ZINC15 database, processed with LigPrep at pH 7.4±0.5 [5].
- Hierarchical Docking: High-throughput virtual screening (HTVS) followed by standard precision (SP) and extra precision (XP) docking modes in Glide [5].
- Binding Affinity Assessment: MM-GBSA (Molecular Mechanics Generalized Born Surface Area) calculations to determine binding free energies [5].
- Stability Validation: Molecular dynamics simulations (100+ ns) to assess compound stability in the binding pocket [5] [8].
Table 2: Key Research Reagent Solutions for STAT3-SH2 Domain Studies
| Reagent/Resource | Specifications | Research Application | Key Features |
|---|---|---|---|
| STAT3 SH2 Domain Structure | PDB ID: 1BG1 (core STAT3 dimer); 6NJS (higher resolution) | Molecular docking and dynamics | Source of 3D structural information for computational studies |
| Compound Libraries | SPECS (~110,000 compounds); ZINC15 natural products (~180,000 compounds) | Virtual screening | Diverse chemical space for hit identification |
| Molecular Dynamics Software | GROMACS; Desmond | Simulation of domain flexibility | Analyzes conformational dynamics and binding stability |
| Docking Algorithms | Glide (HTVS, SP, XP modes); Induced Fit Docking | Virtual ligand screening | Predicts binding poses and affinities |
| Cell-Based Assay Systems | MDA-MB-231, MDA-MB-468 breast cancer lines; Kasumi-1 AML line | In vitro validation | Models for testing STAT3 inhibition efficacy |
Therapeutic Targeting Strategies
Evolution of STAT3 SH2 Domain Inhibitors
The development of STAT3 inhibitors has progressed through several generations, each addressing limitations of previous approaches:
- First-Generation Peptide Inhibitors: Phosphotyrosylated peptides based on STAT3 binding motifs (e.g., pY905LPQTV from gp130). While demonstrating high affinity, these compounds suffered from proteolytic instability, poor oral bioavailability, and limited membrane permeability [3] [4].
- Peptidomimetics: Conformationally constrained compounds like CJ-887 (Kᵢ = 15 nM) offered improved binding affinity but retained poor drug-like properties and cell permeability [3] [4].
- Early Small-Molecule Inhibitors: Initial SB-VLS efforts identified compounds with favorable drug-like properties but often with weak binding affinities, potentially due to neglecting domain flexibility [2] [3].
- Advanced Small-Molecule Inhibitors: Recent approaches incorporating MD simulations have identified neutral, low-molecular-weight compounds with improved potency and drug-like properties [2] [3] [4].
Promising Inhibitor Classes
Recent studies have identified several promising inhibitor classes targeting the STAT3 SH2 domain:
- Neutral Small Molecules: Kong et al. identified two highly potent, neutral, low-molecular-weight STAT3 inhibitors with favorable drug-like properties that directly target the pY+0 pocket without negatively charged moieties [2] [3] [4].
- Natural Product-Derived Inhibitors: Recent screening efforts identified (−)-Epigallocatechin gallate, Kaempferol-3-O-rutinoside, and Saikosaponin D as natural compounds with significant STAT3-SH2 inhibitory potential and favorable ADME/tox profiles [5] [8].
- Specificity-Enhanced Compounds: Modern design strategies focus on compounds that specifically disrupt STAT3 dimerization while minimizing interference with mitochondrial STAT3 functions to reduce adverse effects [3] [4].
The canonical αβββα sandwich architecture of the STAT3 SH2 domain and its specialized pY+0 binding pocket represent a sophisticated structural framework for specific phosphotyrosine recognition and a promising target for therapeutic intervention. The integration of molecular dynamics simulations with advanced structural biology and computational screening methods has dramatically improved our understanding of SH2 domain flexibility and its implications for drug discovery. By accounting for the dynamic nature of this domain and employing sophisticated screening methodologies, researchers have identified novel inhibitor classes with improved potency, specificity, and drug-like properties. Continuing advances in our understanding of STAT3 SH2 domain dynamics, combined with innovative targeting strategies, hold significant promise for developing effective therapeutics for STAT3-driven cancers and other diseases.
The Src Homology 2 (SH2) domain represents a critical modular unit within metazoan signaling pathways, functioning as a specialized reader of phosphotyrosine (pY) motifs to orchestrate protein-protein interactions in signal transduction networks [9]. Within the STAT (Signal Transducer and Activator of Transcription) family of transcription factors, the SH2 domain transcends its conventional adaptor role to become indispensable for multiple facets of molecular activation, including receptor recruitment, phosphorylation-dependent activation, and the critical dimerization that enables nuclear translocation and DNA binding [10] [11]. The uniqueness of the STAT-type SH2 domain is not merely academic; it represents a structural and functional adaptation that has become a focal point for therapeutic intervention in diseases ranging from cancer to immunological disorders [5]. This technical guide delineates the key distinguishing characteristics of STAT-type SH2 domains from the more conventional Src-type SH2 domains, frames these differences within the context of molecular dynamics and flexibility research, and provides methodologies essential for researchers investigating this critical protein domain.
Structural Divergence: A Tale of Two Folds
Despite a conserved core fold, STAT-type and Src-type SH2 domains exhibit significant structural variations that directly impact their function and druggability. All SH2 domains share a fundamental αβββα motif—a central anti-parallel β-sheet (βB-βD) flanked by two α-helices (αA and αB) [10] [9]. This core structure creates two primary subpockets: the pY pocket for phosphotyrosine binding and the pY+3 pocket that confers binding specificity [10]. The critical structural divergence emerges in the C-terminal region following this core motif.
Table 1: Fundamental Structural Classification of SH2 Domains
| Feature | STAT-Type SH2 Domain | Src-Type SH2 Domain |
|---|---|---|
| Core Structure | αA-βB-βC-βD-αB (αβββα motif) | αA-βB-βC-βD-αB (αβββα motif) |
| C-terminal Region | Contains an additional α-helix (αB') | Contains extra β-sheets (βE and βF) |
| Representative Proteins | STAT1, STAT2, STAT3, STAT4, STAT5A, STAT5B, STAT6 | SRC, ABL1, FYN, LCK, ZAP70, SYK, GRB2 |
| Primary Function | Mediates receptor recruitment & STAT dimerization | Facilitates protein relocalization & complex assembly |
The STAT-type SH2 domain is characterized by the presence of an additional α-helix (αB') in this C-terminal region, often referred to as the evolutionary active region (EAR) [10] [6]. Conversely, the Src-type SH2 domain harbors extra β-sheets (βE and βF) instead of this helix [9]. This disparity is not merely structural decoration; it reflects an evolutionary adaptation. Evidence suggests that the linker-SH2 domain of STAT is one of the most ancient and fully developed functional domains, serving as an evolutionary template for the SH2 domain itself [6]. This domain has been identified in plants, suggesting it predates the divergence of plants and animals, while Src-type domains appeared later in metazoan evolution [6].
Figure 1: Structural classification of SH2 domains, highlighting the shared core αβββα motif and the distinctive C-terminal structural elements that define STAT-type and Src-type subgroups.
Functional Specialization: From Dimerization to Disease
The structural uniqueness of the STAT-SH2 domain directly enables its specialized functional capabilities, particularly in mediating STAT dimerization—a process critical for its role as a transcription factor.
Mechanism of STAT Activation and Dimerization
The STAT-SH2 domain orchestrates a multi-step activation process. Initially, it facilitates the recruitment of STAT proteins to phosphorylated tyrosine motifs on activated cytokine receptors [12] [11]. Following receptor recruitment, STATs are phosphorylated by Janus kinases (JAKs) or receptor kinases at a conserved C-terminal tyrosine residue. This phosphorylation triggers a profound conformational change: the SH2 domain of one STAT monomer engages the phosphorylated tyrosine (pY) of another, forming a functional dimer that translocates to the nucleus to drive transcription [10] [13]. This SH2-pY interaction is therefore the linchpin for activated STAT dimer formation. Research on Stat1 and Stat2 has demonstrated that their SH2 domains mediate multiple interactions, including both homo- and heterodimerization, providing evidence that a single SH2-phosphotyrosyl interaction is sufficient for this process [11].
Impact of Mutations on SH2 Domain Function
The functional criticality of the STAT-SH2 domain is underscored by its status as a mutational hotspot in human disease. Sequencing of patient samples has revealed numerous point mutations within the SH2 domains of STAT3 and STAT5B, which can have either gain-of-function (GOF) or loss-of-function (LOF) consequences [10] [14].
Table 2: Functional Consequences of Select STAT-SH2 Domain Mutations
| STAT Protein | Mutation | Location/Region | Pathological Association | Functional Type |
|---|---|---|---|---|
| STAT3 | S614R | BC Loop / pY Pocket | T-LGLL, NK-LGLL, ALK-ALCL | Activating (GOF) [10] |
| STAT3 | K591E, K591M | αA Helix / pY Pocket | AD-HIES | Inactivating (LOF) [10] |
| STAT5B | Y665F | SH2 Domain Interface | T-LGLL, T-PLL | Activating (GOF) [14] |
| STAT5B | Y665H | SH2 Domain Interface | T-PLL (Single Case) | Loss-of-Function (LOF) [14] |
For instance, the STAT5BY665F mutation, a recurrent finding in T-cell leukemias, exemplifies a GOF mutation. In silico modeling predicted that this mutation stabilizes the SH2 domain structure, potentially by promoting intramolecular aromatic stacking interactions [14]. This was confirmed in primary T-cells and mouse models, where the Y665F variant showed enhanced STAT5 phosphorylation, DNA binding, and transcriptional activity [14]. In contrast, the STAT5BY665H mutation at the same residue introduces a histidine imidazole group, predicted to destabilize intramolecular interactions and demonstrated to result in LOF characteristics, including diminished T-cell populations [14]. This illustrates the delicate structural balance within the SH2 domain, where single amino acid changes can fundamentally alter STAT function and lead to divergent disease states.
Dynamics and Flexibility: Implications for Drug Discovery
The conformational plasticity of STAT-SH2 domains presents both challenges and opportunities for therapeutic targeting. Molecular dynamics (MD) simulations have been instrumental in revealing that STAT SH2 domains exhibit significant flexibility, even on sub-microsecond timescales [10] [13].
Key Methodological Approaches for Studying STAT-SH2 Dynamics
Molecular Dynamics (MD) Simulations:
- System Setup: Begin with high-resolution crystal structures (e.g., PDB: 1BG1 for STAT3, 1BF5 for STAT1). Missing loops and residues can be modeled using tools like Modeller [13].
- Simulation Parameters: Perform simulations in explicit water using packages like Desmond, GROMACS, or NAMD. Apply periodic boundary conditions and particle mesh Ewald electrostatics. Maintain constant temperature and pressure (e.g., NPT ensemble) [13].
- Trajectory Analysis: Calculate root mean square deviation (RMSD) and fluctuation (RMSF) to assess global and local stability. Employ principal component analysis (PCA) and k-means clustering to identify dominant conformational substates and collective motions [13].
Computational Screening for SH2 Domain Inhibitors:
- Protein Preparation: Retrieve STAT3-SH2 structures from the PDB (e.g., 6NJS). Process using protein preparation wizards to add hydrogens, fill missing side chains, and minimize energy using a force field like OPLS3e [5].
- Ligand Docking: Use GLIDE or similar docking software with a grid generated around the co-crystallized ligand's location. Perform multi-stage docking: High-Throughput Virtual Screening (HTVS) → Standard Precision (SP) → Extra Precision (XP) to screen large compound libraries [5].
- Binding Affinity Assessment: Conduct Molecular Mechanics/Generalized Born Surface Area (MM-GBSA) calculations on top-scoring complexes to estimate binding free energy (ΔG˅Binding) [5].
- Validation: Perform WaterMap analysis to determine the role of hydration sites in binding and conduct molecular dynamics simulations (≥100 ns) to assess complex stability [5].
These simulations have shown that the STAT3 dimer undergoes a significant "scissor-like" conformational change when bound to DNA, a motion not observed to the same extent in the Stat1 dimer [13]. This large-scale domain motion is driven by more favorable DNA-protein interaction energies and results in a tightening of the SH2 domains. Crucially, during these dynamics, water molecules can diffuse into cavities beneath the dimer interface, expanding pre-existing pockets that could serve as potential binding sites for allosteric inhibitors [13]. This highlights the importance of accounting for protein flexibility and solvation in STAT-directed drug discovery, as crystal structures may not capture all accessible, targetable states [10].
Figure 2: A representative workflow for computational analysis of STAT-SH2 domain dynamics and inhibitor screening, integrating molecular dynamics and virtual screening protocols.
Research Toolkit: Essential Reagents and Methodologies
The following toolkit compiles key reagents and methodological solutions employed in contemporary STAT-SH2 domain research, as derived from cited experimental and computational studies.
Table 3: Research Reagent Solutions for STAT-SH2 Domain Investigation
| Reagent / Solution | Specifications / Function | Experimental Context |
|---|---|---|
| STAT3 Crystal Structure | PDB ID: 6NJS (Resolution: 2.70 Å); used for docking/MD studies due to lack of SH2 domain mutations. | Computational docking & dynamics [5] |
| Natural Compound Library | 182,455 compounds from ZINC15 database; source of potential SH2 domain inhibitors. | Virtual screening for STAT3-SH2 inhibitors [5] |
| Schrödinger Maestro Suite | Software suite (version 2024-2); includes GLIDE, Desmond, Prime for docking, MD, and MM-GBSA. | Integrated computational drug discovery [5] |
| OPLS3e Force Field | Optimized Potential for Liquid Simulations; used for protein/ligand energy minimization and MD. | Protein preparation & molecular dynamics [5] |
| AlphaFold3 & COORDinator | Neural network-based tools for protein structure prediction and mutation energy impact analysis. | Predicting structural & functional impact of SH2 mutations (e.g., STAT5B-Y665F/H) [14] |
| Pathogenicity Prediction Tools | AlphaMissense, CADD, REVEL; computational assessment of mutation pathogenicity. | Classifying STAT-SH2 domain variants [14] |
The STAT-type SH2 domain is a structurally and functionally distinct variant of the canonical SH2 fold, characterized by its unique C-terminal αB' helix and its specialized, essential role in mediating STAT dimerization for transcriptional activation. Its pronounced conformational flexibility, revealed through molecular dynamics simulations, and its status as a mutational hotspot in diseases like cancer and immunodeficiency, underscore its biological and clinical significance. The ongoing structural and dynamic characterization of this domain, facilitated by the experimental and computational methodologies detailed in this guide, continues to illuminate the mechanisms of STAT signaling and uncover novel, targetable pockets for therapeutic intervention. Future research leveraging advanced biophysical techniques and dynamic structural models will be crucial for translating this knowledge into effective targeted therapies.
The molecular flexibility of Src Homology 2 (SH2) domains, particularly those within the Signal Transducer and Activator of Transcription (STAT) family, represents a critical frontier in understanding cellular signaling dynamics and developing targeted therapeutic interventions. As specialized protein modules that recognize phosphorylated tyrosine (pTyr) motifs, SH2 domains mediate precise protein-protein interactions that drive fundamental processes including cell proliferation, differentiation, and immune responses [15] [16]. The STAT family of transcription factors exemplifies the crucial role of SH2 domains in signal transduction, where their flexibility and conformational dynamics govern dimerization, nuclear translocation, and gene expression [9] [1]. Within the broader context of molecular dynamics and STAT SH2 domain research, this technical guide examines the structural elements that confer flexibility—specifically key residues and dynamic loops—and their implications for function and dysfunction in human disease. Through integrated experimental and computational approaches, researchers are unraveling how these molecular determinants enable STAT SH2 domains to serve as dynamic regulators within complex cellular networks, providing insights for targeting pathological signaling in cancer and other disorders.
Structural Architecture of SH2 Domains
SH2 domains constitute a conserved structural fold of approximately 100 amino acids that specifically recognizes pTyr-containing sequences [16] [9]. The canonical SH2 domain structure adopts a sandwich-like architecture composed of a central antiparallel β-sheet flanked by two α-helices, designated as αA and αB [16] [9]. This core scaffold maintains remarkable conservation across the human SH2 domain family, which encompasses approximately 110 proteins containing 120 distinct SH2 domains [16] [1].
The phosphotyrosine-binding pocket represents the most conserved structural feature, characterized by a critical arginine residue (βB5) within the highly conserved FLVR motif that forms salt bridges with the phosphate moiety of pTyr [16] [9]. Beyond this universal pTyr recognition capability, SH2 domains exhibit considerable specificity for residues C-terminal to the pTyr, primarily determined by structural variations in loops and secondary binding pockets [17]. These variable regions enable different SH2 domains to recognize distinct sequence motifs, thereby conferring specificity in signaling pathways.
Table 1: Core Structural Elements of SH2 Domains
| Structural Element | Description | Functional Role |
|---|---|---|
| Central β-sheet | 3-7 antiparallel β-strands | Forms structural core and binding surface |
| αA and αB helices | Flank central β-sheet | Provide structural stability |
| pTyr-binding pocket | Contains conserved Arg (βB5) | Recognizes phosphate moiety of pTyr |
| Specificity pockets | Adjacent to pTyr pocket | Bind residues C-terminal to pTyr (P+1 to P+4) |
| Connecting loops | Variable length sequences | Control access to binding pockets |
STAT-type SH2 domains exhibit distinctive structural adaptations that differentiate them from SRC-type SH2 domains [9]. Specifically, STAT SH2 domains lack the βE and βF strands present in most other SH2 domains and feature a split αB helix [9]. This structural modification likely facilitates the domain-swapped dimerization critical for STAT activation and nuclear function [9]. Additionally, STAT SH2 domains possess more open binding surfaces due to reduced loop obstruction, which may accommodate their specific recognition of pYxxQ motifs (where x represents any amino acid) [17].
Key Determinants of SH2 Domain Flexibility and Specificity
Dynamic Loops as Specificity Gates
The flexible loops connecting secondary structural elements play a pivotal role in governing SH2 domain specificity by controlling access to binding pockets. Research has revealed that loops function as molecular gates that either permit or restrict ligand access to specificity-determining pockets [17]. This gating mechanism explains how diverse binding specificities can emerge from a conserved structural scaffold.
The EF loop (connecting β-strands E and F) and BG loop (connecting α-helix B and β-strand G) constitute particularly important structural elements that define the shape and accessibility of binding pockets [9] [17]. In many SH2 domains, these loops form a hydrophobic cavity that recognizes residues at the P+3 position relative to the pTyr [17]. However, in SH2 domains with different specificities, these loops may physically block certain pockets while permitting access to others. For instance, in Grb2's SH2 domain, which recognizes pYxN motifs, a bulky tryptophan residue in the EF loop occupies the P+3 binding pocket, forcing the bound peptide to adopt a β-turn conformation and enabling specific recognition of asparagine at P+2 [17].
Table 2: Loop-Mediated Specificity Determinants in SH2 Domains
| SH2 Domain Group | Recognized Motif | Key Loop Determinants | Structural Consequence |
|---|---|---|---|
| Group IA/IB | pYxxψ (ψ = hydrophobic) | Open EF/BG loops | Forms accessible P+3 hydrophobic pocket |
| Group IC | pYxN | Bulky EF1 residue (Trp) | Blocks P+3 pocket, enables P+2 Asn recognition |
| Group IIC | pYxxxψ | Open P+4 pocket | BG loop residue displacement creates P+4 pocket |
| STAT-type | pYxxQ | Reduced loop obstruction | Open binding surface for dimerization |
The BRDG1/STAP-1 SH2 domain exemplifies an extreme case of loop-mediated specificity, where structural analyses revealed a unique hydrophobic pocket that accommodates residues at the P+4 position [17]. This "pentagon basket" pocket is formed by five hydrophobic residues and is inaccessible in most other SH2 domains because it is occupied by a leucine or isoleucine side chain from the BG loop [17]. In BRDG1, alternative BG loop sequences leave this pocket open, enabling recognition of P+4 hydrophobic residues and demonstrating how loop variations dramatically alter binding specificity.
Key Residues Governing Binding Energetics
Beyond structural loops, specific residues critically influence SH2 domain flexibility and function through their roles in binding energetics and conformational stability. The highly conserved arginine residue (βB5) in the FLVR motif is absolutely essential for pTyr recognition, forming direct salt bridges with the phosphate moiety [15] [16]. Mutation of this residue typically abolishes phosphopeptide binding, underscoring its fundamental importance.
The specificity of SH2 domain-phosphopeptide interactions is characterized by moderate binding affinities (Kd values typically ranging from 0.1–10 μM) that allow for specific yet reversible interactions necessary for dynamic signaling processes [9] [1]. These affinities are determined by the composite energetics of residues surrounding the pTyr. Quantitative analyses using bacterial surface display and deep sequencing have revealed that the free energy of binding (ΔG) depends on specific amino acids at positions P+1 to P+4 C-terminal to the phosphotyrosine [18]. For example, the c-Src SH2 domain preferentially binds pYEEI motifs, with glutamic acid residues at P+1 and P+2 contributing favorably to binding energetics, while an isoleucine at P+3 provides hydrophobic stabilization [15] [18].
Recent high-throughput studies employing fully randomized peptide libraries and quantitative modeling have enabled precise determination of the energetic contributions of individual residue positions to SH2 domain binding [18]. These approaches demonstrate that binding free energy parameters (ΔΔG/RT) provide more robust and library-independent measures of specificity compared to simple enrichment metrics, allowing accurate prediction of SH2 binding affinities across theoretical sequence space [18].
Experimental Approaches for Analyzing SH2 Flexibility
High-Throughput Specificity Profiling
Comprehensive analysis of SH2 domain flexibility and binding specificity requires experimental approaches that quantitatively measure interactions across vast sequence spaces. Bacterial surface display of peptide libraries coupled with deep sequencing has emerged as a powerful methodology for profiling SH2 domain specificities [18]. This technique involves displaying genetically-encoded peptide libraries on bacterial surfaces, phosphorylating tyrosine residues using kinase domains, and selecting for SH2 domain binding through fluorescence-activated cell sorting or affinity purification.
The experimental workflow typically employs one of two library designs: (1) the "X5YX5" library with a fixed central tyrosine flanked by five degenerate amino acid positions on each side, or (2) fully randomized "X11" libraries where all 11 consecutive positions are variable [18]. Following enzymatic phosphorylation, the library undergoes one or more rounds of selection with purified SH2 domains. Deep sequencing of pre- and post-selection populations enables quantitative assessment of sequence enrichment, which can be modeled to determine binding free energy parameters [18].
Advanced computational frameworks, such as the ProBound algorithm, employ maximum likelihood estimation to model selection data and infer free-energy matrices that predict binding affinity for any peptide sequence within the theoretical space covered by the library [18]. These models account for multiple binding registers and non-specific binding, providing robust, library-independent estimates of the energetic effects of amino acid substitutions [18].
Diagram 1: Workflow for SH2 specificity profiling
Structural and Computational Analysis Methods
Molecular dynamics (MD) simulations provide atomic-level insights into SH2 domain flexibility and conformational dynamics. Several specialized tools enable comprehensive analysis of MD trajectories:
Table 3: Molecular Dynamics Analysis Tools for SH2 Domain Studies
| Tool | Primary Function | Application to SH2 Domains |
|---|---|---|
| MDAnalysis | Flexible trajectory analysis | Analyzing binding interface dynamics |
| MDTraj | Fast trajectory analysis | Calculating RMSD and binding pocket fluctuations |
| VMD | Visualization and analysis | Visualizing loop conformations and binding events |
| CPPTRAJ | Advanced trajectory processing | Time-resolved analysis of domain flexibility |
| PLUMED | Enhanced sampling and free energy calculations | Determining binding energetics and conformational landscapes |
| gmmpbsa/gmxMMPBSA | Binding free energy calculations | Quantifying SH2-phosphopeptide interaction energies |
The CoDIAC (Comprehensive Domain Interface Analysis of Contacts) pipeline represents a specialized framework for structural analysis of SH2 domains [19]. This Python-based package extracts and analyzes contact maps from experimental structures (PDB) and predicted models (AlphaFold) to map interaction interfaces at residue-level resolution [19]. CoDIAC integrates multiple data sources, including PTM databases and genetic variants, to contextualize structural findings with biological annotations. For SH2 domains, this approach has revealed coordinated regulation of binding interfaces by serine/threonine phosphorylation and acetylation, suggesting cross-talk between signaling systems [19].
Research Reagents and Methodologies
Table 4: Essential Research Reagents for SH2 Domain Flexibility Studies
| Reagent/Tool | Specifications | Experimental Application |
|---|---|---|
| SH2 Domain Constructs | Recombinant proteins (wild-type and mutants) | Binding assays, structural studies, specificity profiling |
| Phosphopeptide Libraries | X5YX5 (theoretical diversity: ~10^13) or fully randomized X11 libraries | High-throughput specificity profiling using display technologies |
| Bacterial Display System | Plasmid-encoded peptide display | Library selection and enrichment analysis |
| Tyrosine Kinase Domains | Active kinase domains (e.g., Src, Abl) | Enzymatic phosphorylation of displayed peptide libraries |
| ProBound Software | Statistical learning algorithm | Quantitative modeling of binding free energies from selection data |
| CoDIAC Pipeline | Python-based structural analysis | Comprehensive contact mapping and interface analysis |
| MD Simulation Software | GROMACS, AMBER, NAMD | Atomic-level simulation of conformational dynamics |
Implications for STAT SH2 Domain Research and Therapeutic Targeting
The molecular determinants of STAT SH2 domain flexibility have direct implications for understanding pathological signaling and developing targeted therapies. STAT proteins, particularly STAT3 and STAT5, are frequently hyperactivated in cancers and inflammatory diseases, driving aberrant gene expression programs [9] [1]. Their SH2 domains mediate critical dimerization steps through reciprocal pTyr-SH2 interactions, making them attractive therapeutic targets [9] [1].
The unique structural features of STAT SH2 domains—including their open binding surfaces and adapted loop architectures—create opportunities for selective inhibition [9] [17]. Small molecules that target the SH2 domain and disrupt STAT dimerization have shown promise in preclinical models, though achieving selectivity remains challenging due to conservation of the pTyr-binding pocket [16] [9]. Alternative strategies include targeting allosteric sites or interfacial inhibitors that exploit the dynamic nature of SH2 domains during dimerization [1].
Emerging research also highlights non-canonical functions of SH2 domains beyond simple pTyr recognition. Many SH2 domains, including those in STAT proteins, interact with membrane phospholipids such as phosphatidylinositol-4,5-bisphosphate (PIP2) and phosphatidylinositol-3,4,5-trisphosphate (PIP3) [16] [9]. These interactions often involve cationic regions near the pTyr-binding pocket and can modulate membrane localization and signaling output [16]. Additionally, SH2 domains participate in liquid-liquid phase separation (LLPS) through multivalent interactions, forming biomolecular condensates that enhance signaling efficiency [16] [9]. In T-cell receptor signaling, interactions between GRB2, Gads, and LAT receptors undergo phase separation that enhances signaling capacity [16]. Similar mechanisms may operate in STAT signaling pathways, where multivalency and post-translational modifications could drive condensate formation with functional consequences for gene regulation.
Diagram 2: STAT activation pathway and disease linkage
Understanding the flexibility determinants of STAT SH2 domains thus provides a multidimensional perspective on their function, encompassing atomic-level interactions, conformational dynamics, higher-order assembly, and pathological misregulation. This integrated view continues to inspire novel therapeutic approaches that target these critical signaling modules in human disease.
The Src Homology 2 (SH2) domain, a module of approximately 100 amino acids, has been fundamentally understood for decades as a phosphotyrosine (pY) binding unit that directs the assembly of signaling complexes in protein tyrosine kinase (PTK) pathways [20] [21]. However, emerging research reveals a functional landscape for SH2 domains that extends far beyond this canonical role. It is now evident that SH2 domains participate in lipid interactions and facilitate the formation of biomolecular condensates through liquid-liquid phase separation (LLPS), processes critical for spatiotemporal control of cellular signaling [9] [16] [22]. This expanded understanding is particularly relevant for STAT (Signal Transducer and Activator of Transcription) proteins, whose SH2 domains are essential for dimerization, nuclear translocation, and transcriptional activity [23]. The molecular dynamics and flexibility of STAT SH2 domains underpin their ability to engage in these diverse interactions, making them a focal point for therapeutic intervention. This review synthesizes recent advances that redefine SH2 domains as versatile regulatory modules, framing these discoveries within the context of STAT SH2 domain research and drug development.
Canonical Structure and the Foundation for Novel Functions
Conserved Architecture and pY-Peptide Recognition
The classic SH2 domain fold consists of a central antiparallel β-sheet flanked by two α-helices, forming a βαββββαβ structure [9] [23] [21]. This scaffold creates two principal ligand-binding sites: a highly conserved pY-binding pocket and a more variable specificity-determining region. The pY-binding pocket, located within the βB strand, contains a critical arginine residue (βB5) that forms a salt bridge with the phosphate moiety of the phosphotyrosine ligand [9] [16]. The specificity of individual SH2 domains is conferred by residues that interact with amino acids C-terminal to the pY, typically at the pY+1 to pY+5 positions [21]. In STAT proteins specifically, the SH2 domain is essential for reciprocal phosphotyrosine-mediated dimerization, which is a prerequisite for their nuclear translocation and function as transcription factors [23].
Molecular Dynamics and Plasticity
The structural conservation of SH2 domains belies a significant degree of conformational flexibility. This plasticity enables certain SH2 domains to recognize diverse ligands, including those without phosphotyrosine, such as serine/threonine-phosphorylated sequences, phosphatidylinositol lipids, and even unphosphorylated motifs [21]. This adaptability is governed by the thermodynamic and kinetic properties of the domains, which allow for rapid cellular responses to changing conditions [20]. The molecular dynamics of SH2 domains, including loop flexibility and side-chain rearrangements, are fundamental to their emerging roles in lipid binding and phase separation, as these processes often require multivalent, low-affinity interactions that are highly sensitive to the cellular environment.
Emerging Non-Canonical Functions of SH2 Domains
SH2 Domains as Lipid-Binding Modules
Recent studies have revealed that a significant proportion of SH2 domains interact with membrane lipids, expanding their function beyond soluble protein-protein interactions. Table 1 summarizes key SH2-containing proteins with demonstrated lipid-binding activity and their functional roles.
Table 1: Lipid-Binding Capabilities of SH2 Domain-Containing Proteins
| Protein Name | Function of Lipid Association | Lipid Moiety | Biological Role |
|---|---|---|---|
| SYK | PIP3-dependent membrane binding required for non-catalytic activation of STAT3/5 [16]. | PIP3 | Scaffolding function in immune signaling. |
| ZAP70 | Facilitates and sustains interactions with TCR-ζ chain [16]. | PIP3 | T-cell receptor signaling. |
| LCK | Modulates interaction with binding partners in the TCR signaling complex [16]. | PIP2, PIP3 | Early T-cell activation. |
| ABL | Membrane recruitment and modulation of Abl kinase activity [16]. | PIP2 | Regulation of cytoskeletal dynamics. |
| VAV2 | Modulates interaction with membrane receptors (e.g., EphA2) [16]. | PIP2, PIP3 | Guanine nucleotide exchange factor (GEF) activity. |
| C1-Ten/Tensin2 | Regulates Abl activity and IRS-1 phosphorylation in insulin signaling [9] [16]. | PIP3 | Insulin signaling pathway. |
The mechanistic basis for lipid recognition often involves cationic regions near the pY-binding pocket, which are typically flanked by aromatic or hydrophobic side chains [9] [16]. This structural arrangement allows the domain to interact with negatively charged phospholipid head groups, such as phosphatidylinositol-4,5-bisphosphate (PIP2) and phosphatidylinositol-3,4,5-trisphosphate (PIP3). From a functional perspective, lipid binding serves to recruit SH2-containing proteins to the plasma membrane, dramatically increasing their local concentration and facilitating encounters with phosphorylated receptor targets. This membrane recruitment can also allosterically modulate enzymatic activity or scaffolding function, as demonstrated in the cases of SYK, VAV, and ZAP70 [9] [16]. Furthermore, mutations within these lipid-binding pockets have been linked to human disease, underscoring their physiological importance and highlighting a new avenue for therapeutic targeting, such as the development of nonlipidic inhibitors for SYK kinase [9] [16].
SH2 Domains in Biomolecular Condensate Formation via Phase Separation
Liquid-liquid phase separation (LLPS) has emerged as a fundamental mechanism for cellular organization, and SH2 domain-containing proteins are prominent players in this process. Their ability to engage in multivalent interactions—both through their SH2 domains and other modular domains like SH3—makes them ideal drivers of condensate assembly [9] [22]. Table 2 provides examples of signaling condensates where SH2 domain-mediated interactions are crucial.
Table 2: SH2 Domain-Containing Proteins in Biomolecular Condensates
| Condensate Complex | Biological Role | Key SH2-Containing Proteins | Reference |
|---|---|---|---|
| LAT-GRB2-SOS1 | T-cell receptor activation and signaling amplification. | GRB2, PLCγ1, ZAP70, LCK [16] | |
| FGFR2:SHP2:PLCγ1 | Enhances activity of Receptor Tyrosine Kinase (RTK) signaling. | SHP2, PLCγ1 [16] | |
| N-WASP–NCK | Promotes actin polymerization in podocyte kidney cells and T-cell signaling. | NCK [9] [16] | |
| SLP65, CIN85 | B-cell receptor signaling. | SLP65 [16] | |
| Mutant SHP2 Condensates | Pathological activation of RAS-MAPK signaling in developmental disorders. | SHP2 (NS/JMML and NS-ML mutants) [22] |
A paradigmatic example of the pathological consequences of aberrant phase separation is found in the phosphatase SHP2. Disease-associated mutations in SHP2, found in Noonan syndrome (NS), juvenile myelomonocytic leukemia (JMML), and Noonan syndrome with multiple lentigines (NS-ML), lead to a gain-of-function ability to undergo LLPS [22]. Remarkably, both activating (NS/JMML) and inactivating (NS-ML) mutations result in similar puncta formation and clinical manifestations. This phenomenon is explained by a model where mutant SHP2 proteins form condensates that recruit and hyperactivate wild-type SHP2, leading to sustained RAS-MAPK signaling [22]. The process is driven by the conserved, well-folded PTP domain through multivalent electrostatic interactions and is regulated by an autoinhibitory mechanism involving the N-SH2 domain [22]. This discovery directly links dysregulated LLPS to the pathogenesis of human developmental disorders and cancers.
The following diagram illustrates the sequence of events in mutant SHP2-induced pathological condensate formation and signaling activation:
Figure 1: Pathological Condensate Formation by Mutant SHP2.
Integration of Lipid and Phase Separation Biology
The interactions between lipid membranes and biomolecular condensates represent a frontier in understanding SH2 domain function. Lipid membranes can serve as nucleation platforms for condensate formation, reducing the critical concentration required for phase separation by orders of magnitude—from micromolar to nanomolar levels—through membrane anchoring and thermodynamic coupling [24]. This creates specialized microenvironments that substantially enhance enzymatic activities and signaling output. For instance, phosphotyrosine-driven protein condensation can couple with membrane lipid phase transitions, creating highly organized and efficient signaling platforms [24]. The coupling is regulated by post-translational modifications (e.g., phosphorylation), membrane composition (e.g., cholesterol content), and environmental factors (e.g., calcium ions) [24]. This integrated view positions SH2 domains at the nexus of protein-protein, protein-lipid, and phase separation events, orchestrating the precise spatiotemporal dynamics of cellular signaling networks.
Experimental Approaches for Investigating Novel SH2 Functions
Methodologies for Studying Lipid Interactions
- Hydrogen–Deuterium Exchange Mass Spectrometry (HDX-MS): This technique probes protein dynamics and membrane interactions by measuring the exchange rate of backbone amide hydrogens with deuterium in the solvent. It has been successfully used to identify intramolecular contacts, such as those between the SH2 and C2 domains in SHIP1, that regulate membrane localization and autoinhibition [25].
- Single-Molecule Measurements on Supported Lipid Bilayers (SLBs): Purified proteins are introduced onto artificial lipid bilayers, and their binding frequency, dwell time, and diffusion are observed using Total Internal Reflection Fluorescence Microscopy (TIRF-M). This approach directly visualizes how the SH2 domain of SHIP1 autoinhibits membrane binding and how this block is relieved by phosphotyrosine ligands [25].
- In Vitro Reconstitution and Binding Assays: These involve testing the binding of purified SH2 domains or full-length proteins to lipid vesicles of defined composition. This allows for the quantitative assessment of lipid-binding specificity and affinity.
Techniques for Probing Phase Separation
- Live-Cell Fluorescence Microscopy: The foundational technique for observing puncta formation in cells. Proteins of interest are tagged with fluorescent proteins (e.g., mEGFP, mScarlet) and expressed in relevant cell lines. High-content image analysis can quantify puncta number, size, and density [22].
- Fluorescence Recovery After Photobleaching (FRAP): This method is used to confirm the liquid-like properties of condensates. A region within a condensate is photobleached, and the recovery of fluorescence due to the exchange of molecules with the surrounding solution is monitored over time. Rapid recovery is indicative of a dynamic, liquid-like state [22] [24].
- In Vitro Phase Separation Assays: Recombinant proteins are purified and mixed in physiological buffers to determine the minimal components required for LLPS. Parameters such as protein concentration, salt, and pH can be systematically varied to define the conditions driving phase separation [22].
Computational and In Silico Screening
Computational methods are indispensable for translating mechanistic insights into drug discovery campaigns. For STAT3, a key protein reliant on its SH2 domain for function, in silico screening has been used to identify natural compounds that target the SH2 domain and disrupt STAT3 dimerization [23]. The standard workflow involves:
- Protein and Ligand Preparation: The crystal structure of the STAT3 SH2 domain (e.g., PDB: 6NJS) is prepared, and a library of natural compounds is retrieved from databases like ZINC15.
- Molecular Docking: High-throughput virtual screening (HTVS) is performed, followed by standard precision (SP) and extra precision (XP) docking to predict binding poses and affinities.
- Binding Free Energy Calculations: Molecular Mechanics with Generalized Born and Surface Area Solvation (MM-GBSA) is used to calculate the binding free energy of top hits.
- Molecular Dynamics (MD) Simulations: MD simulations assess the stability of the protein-ligand complex over time, providing insights into conformational dynamics.
- Network Pharmacology: This maps the compound's interactions within biological networks, highlighting potential multi-target effects and off-targets [23].
Figure 2: Computational Screening Workflow for STAT3-SH2 Inhibitors.
Table 3: Essential Research Reagents for Investigating Non-Canonical SH2 Functions
| Reagent / Tool | Function / Application | Example Use Case |
|---|---|---|
| Supported Lipid Bilayers (SLBs) | In vitro reconstitution of cellular membranes to study protein-lipid interactions and binding kinetics. | Measuring SHIP1 membrane binding dynamics [25]. |
| Fluorescent Protein Tags (mEGFP, mScarlet) | Labeling proteins for live-cell imaging and tracking of localization and condensate formation. | Visualizing SHP2 mutant puncta formation in cells [22]. |
| Allosteric SH2 Domain Inhibitors | Small molecules that target the autoinhibitory interface or regulatory sites, modulating protein conformation and activity. | Attenuating LLPS of disease-associated SHP2 mutants [22]. |
| Combinatorial Phosphopeptide Libraries | High-throughput profiling of SH2 domain binding specificity and sequence preferences. | Determining binding motifs for canonical pY-peptide recognition [21]. |
| OPLS3e Force Field | A physics-based model for energy calculations in molecular dynamics simulations and docking studies. | Energy minimization and MM-GBSA calculations for STAT3-SH2 inhibitors [23]. |
| QikProp Tool | Computational prediction of pharmacokinetic properties (ADME) of small molecule hits. | Prioritizing natural compound leads with drug-like properties [23]. |
The paradigm of SH2 domain function has evolved from a static view of pY-peptide recognition to a dynamic model encompassing lipid binding and biomolecular condensate formation. These non-canonical functions are deeply intertwined with the molecular dynamics and conformational flexibility of the domains themselves. For STAT proteins and other SH2-containing signaling molecules, these mechanisms enable rapid, reversible, and spatially constrained activation of downstream pathways. The discovery that disease-associated mutations can cause pathological phase separation, as seen in SHP2, opens a new chapter in understanding the molecular etiology of developmental disorders and cancers. Targeting these emergent properties—such as with allosteric inhibitors that disrupt aberrant LLPS or compounds that block pathological protein-lipid interactions—represents a promising and innovative therapeutic strategy. Future research will undoubtedly focus on quantitatively mapping the interplay between SH2 domain dynamics, membrane environment, and condensate formation, leveraging advanced techniques in structural biology, biophysics, and computation to develop the next generation of targeted therapeutics.
Computational Strategies for Capturing and Exploiting SH2 Dynamics
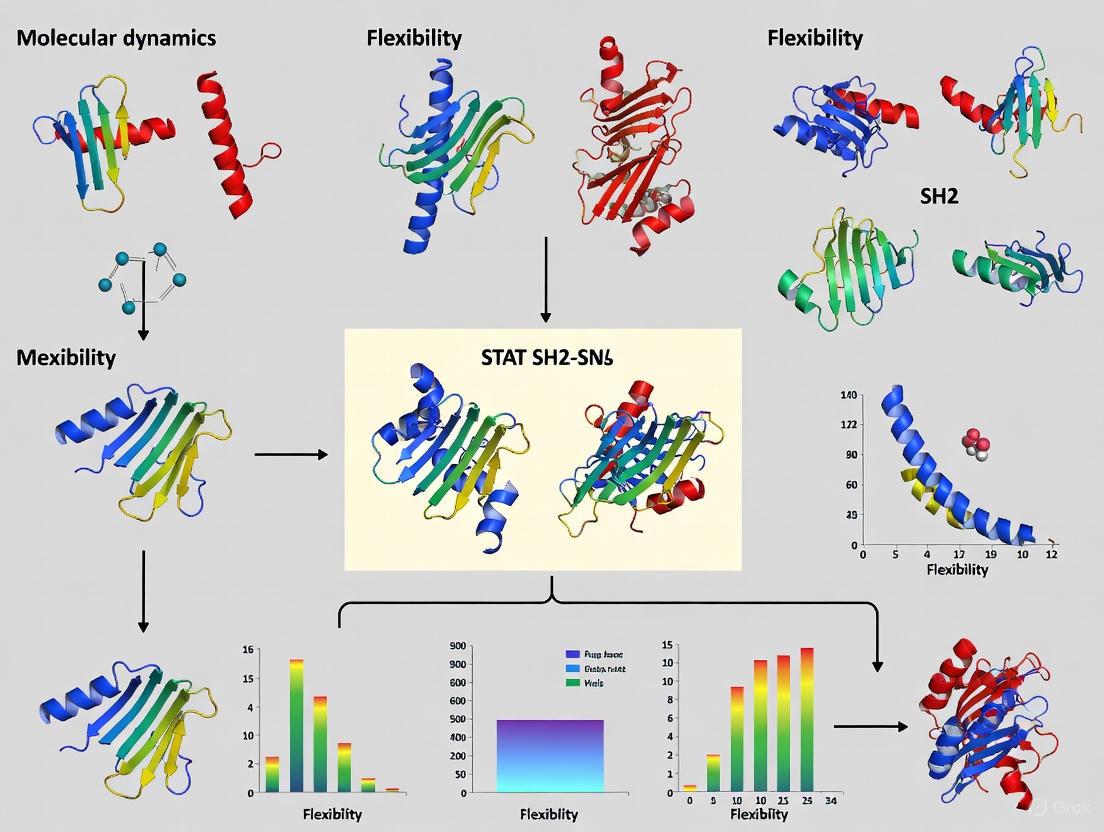
Molecular dynamics (MD) simulations have become an indispensable tool for understanding the behavior of biomolecules at an atomic level, covering timescales from nanoseconds to microseconds [26]. These simulations provide a dynamic view of molecular systems, moving beyond static snapshots to capture the essential motions that govern biological function. Within the context of drug discovery, MD simulations are particularly valuable for studying transcription factors like STAT3, which have historically been considered "undruggable" due to the large size of their protein-protein interaction interfaces [4]. The Src Homology 2 (SH2) domain of STAT3 is a particularly compelling target, as it facilitates the dimerization essential for STAT3's activation and subsequent nuclear translocation [5]. Disrupting this domain offers a promising strategy for cancer therapy, but effective drug design requires a deep understanding of the domain's conformational flexibility—a understanding that MD simulations are uniquely positioned to provide.
STAT3 activation is driven by its SH2 domain, which binds to a phosphorylated tyrosine residue (Y705) of another STAT3 molecule to form an active dimer [5]. This interaction occurs within a binding pocket divided into three sub-pockets: pY+X (hydrophobic side), pY+0 (binds to pY705), and pY+1 (binds to L706) [5]. The structural flexibility of this pocket, particularly its high mobility noted in crystal structures [4], presents both a challenge and an opportunity for inhibitor development. Molecular dynamics simulations enable researchers to capture this flexibility, providing insights that are critical for identifying and optimizing small molecules that can effectively disrupt STAT3 function.
Fundamental Principles of Molecular Dynamics Simulations
Theoretical Foundations
Molecular dynamics simulations operate on the principle of numerically integrating Newton's equations of motion for a system of particles [27]. In classical MD, molecules are represented as collections of atoms or groups of atoms, each assigned parameters for mass, charge, and interactions [27]. The simulation system is propagated through time using deterministic rules, generating a trajectory that describes the system's evolution. This trajectory can then be analyzed to extract structural, dynamic, and thermodynamic properties of the molecular system [27].
The potential energy of the system is described by a force field, which includes terms for bonded interactions (bonds, angles, dihedrals) and non-bonded interactions (electrostatics, van der Waals) [27]. The quality of a simulation heavily depends on the chosen force field and its parameters. For biomolecular systems in condensed phases, molecular mechanics (MM) force fields are typically employed because they offer a balance between computational efficiency and accuracy, allowing simulations of systems containing tens to hundreds of thousands of atoms [27].
Key Methodological Considerations
Several critical methodological choices determine the success and biological relevance of an MD simulation. The simulation system must be constructed to mimic the native environment as closely as possible [26]. This typically involves solvating the protein in water, adding ions to neutralize the system's charge, and applying periodic boundary conditions to minimize edge effects [26]. The choice of integration timestep is constrained by the fastest motions in the system (typically bond vibrations involving hydrogen), often requiring the use of holonomic constraints on these bonds to enable longer timesteps [27].
Proper sampling is essential for obtaining meaningful results, as many properties of interest depend on the correct distribution of states rather than single optimal configurations [27]. For proteins like STAT3, relevant timescales can span from nanoseconds for local sidechain motions to microseconds or longer for larger conformational changes [27]. Modern hardware has made microsecond-length simulations routine for biological systems of 50-100,000 atoms, though herculean efforts have pushed simulations into the millisecond range [27].
Table 1: Key Stages in Molecular Dynamics Simulations
| Stage | Purpose | Key Tools/Commands |
|---|---|---|
| System Setup | Prepare protein structure, define simulation box, solvation | pdb2gmx, editconf, solvate |
| Minimization | Remove steric clashes and high-energy configurations | grompp, genion |
| Equilibration | Gradually bring system to target temperature and pressure | Position restraints, thermostat/barostat |
| Production Run | Generate trajectory for analysis | Long simulation with no restraints |
| Analysis | Extract biologically relevant information from trajectory | RMSD, RMSF, H-bond analysis |
Computational Methodologies for Studying SH2 Domain Flexibility
MD Simulation Protocol for SH2 Domains
The following protocol outlines a general approach for conducting MD simulations of SH2 domains, adapted from established methodologies [26] with specific applications to STAT3 SH2 domains [5] [4]:
Obtain and Prepare Protein Coordinates: Download the STAT3 SH2 domain structure from the Protein Data Bank (e.g., PDB ID 6NJS, chosen for its better resolution and lack of mutations in the SH2 domain) [5]. Preprocess the structure using tools like Schrödinger's Protein Preparation Wizard or GROMACS's
pdb2gmxto add hydrogen atoms, fill missing side chains, assign bond orders, and minimize energy using a force field such as OPLS3e [5] [26].Define System Boundaries and Solvation: Create a simulation box around the protein using
editconfwith periodic boundary conditions. For a cubic box, maintain a minimum distance of 1.0-1.4 nm from the protein periphery [26]. Solvate the system usingsolvateand add ions (e.g., Na+, Cl-) withgenionto neutralize the system's net charge [26].Energy Minimization and Equilibration: Perform energy minimization to remove steric clashes using the
gromppandmdruncommands. Gradually equilibrate the system through restrained dynamics, first with position restraints on protein heavy atoms while relaxing solvent, then without restraints to bring the entire system to the target temperature (typically 310 K) and pressure (1 bar) [26].Production MD Simulation: Conduct an unrestrained production simulation, typically lasting 100 ns to 1 μs depending on the biological process of interest. Use a timestep of 2 fs with constraints applied to bonds involving hydrogen atoms. Save trajectory frames at regular intervals (e.g., every 100 ps) for subsequent analysis [26].
Trajectory Analysis: Analyze the saved trajectory to calculate properties such as root-mean-square deviation (RMSD) for structural stability, root-mean-square fluctuation (RMSF) for residue flexibility, radius of gyration, hydrogen bonding patterns, and distances between key residues [5] [26].
Diagram: Workflow for Molecular Dynamics Simulations of SH2 Domains
Advanced Sampling and Free Energy Calculations
For studying SH2 domain binding events, advanced sampling techniques are often necessary due to the timescales involved. The Molecular Mechanics Generalized Born Surface Area (MM-GBSA) method provides an efficient approach for calculating binding free energies from MD trajectories [5]. This method combines molecular mechanics energy terms with continuum solvation models to estimate the free energy of binding using the equation:
ΔGBinding = ΔGComplex - (ΔGreceptor + ΔGligand)
where ΔGBinding, ΔGreceptor, and ΔGligand denote the total binding energy of the complex, free receptor, and unbound ligand, respectively [5]. More negative values indicate stronger binding. In studies of STAT3 SH2 domain inhibitors, MM-GBSA calculations have identified compounds with binding free energies ranging from -40 to -60 kcal/mol, correlating with their inhibitory potency [5].
Application to STAT SH2 Domain Flexibility and Drug Discovery
Incorporating SH2 Domain Flexibility in Virtual Screening
Traditional structure-based virtual ligand screening (SB-VLS) often treats the target protein as a rigid structure, which can limit the identification of high-affinity binders for flexible domains like STAT3's SH2 domain [4]. To address this limitation, researchers have developed approaches that incorporate domain flexibility through MD simulations. In one innovative study [4]:
- An MD simulation of the STAT3 SH2 domain in complex with a high-affinity peptidomimetic ligand (CJ-887) was conducted
- An averaged structure from the MD trajectory was calculated and optimized
- This "induced-active site" receptor model was used for virtual screening of 110,000 compounds
- Top hits were selected based on interactions with key pY+0 binding pocket residues R609 and S613
This approach identified two highly potent, neutral, low-molecular weight STAT3 inhibitors with favorable drug-like properties, demonstrating the value of incorporating domain flexibility in drug discovery campaigns [4].
Table 2: Key Research Reagents for SH2 Domain Molecular Dynamics Studies
| Reagent/Resource | Function in Research | Application Example |
|---|---|---|
| STAT3 SH2 Domain Structure (6NJS) | High-resolution protein template for simulations | Molecular docking and dynamics simulations [5] |
| GROMACS MD Suite | Open-source software for MD simulations | Simulation of protein dynamics with various force fields [26] |
| Schrödinger Suite | Commercial software for computational drug discovery | Protein preparation, docking, MM-GBSA calculations [5] |
| ZINC15 Database | Public repository of commercially available compounds | Source of natural products for virtual screening [5] |
| OPLS3e Force Field | Empirical potential function for energy calculations | Energy minimization and molecular dynamics [5] |
| CJ-887 Peptidomimetic | High-affinity STAT3 SH2 domain binder | Reference compound for induced-active site modeling [4] |
Case Study: Natural Product Screening Against STAT3 SH2 Domain
A comprehensive in silico screening study exemplifies the application of MD simulations to STAT3 SH2 domain drug discovery [5]. Researchers screened 182,455 natural compounds from the ZINC15 database against the STAT3 SH2 domain using a multi-step approach:
- Molecular Docking: Compounds were docked using high-throughput virtual screening (HTVS), followed by standard precision (SP) and extra precision (XP) docking modes
- Binding Affinity Assessment: MM-GBSA calculations determined binding free energies for top candidates
- Stability Validation: Molecular dynamics simulations (100-200 ns) assessed complex stability and interaction persistence
- Pharmacokinetic Profiling: QikProp tool evaluated drug-like properties of potential hits
This integrated approach identified ZINC67910988 as a particularly promising candidate, demonstrating superior stability in MD simulations and favorable binding characteristics in WaterMap analysis [5]. The compound maintained stable interactions with key SH2 domain residues throughout the simulation timeframe, suggesting strong potential as a STAT3 inhibitor.
Diagram: Virtual Screening Workflow for STAT3 SH2 Domain Inhibitors
Quantitative Analysis of SH2 Domain Simulations
Key Parameters and Performance Metrics
MD simulations of STAT3 SH2 domains have yielded important quantitative insights into domain flexibility and inhibitor binding. Analysis of simulation trajectories provides metrics for assessing system stability and binding interactions:
- Root-mean-square deviation (RMSD): Measures structural stability over time; stable complexes typically show RMSD values below 2-3 Å after equilibration [5]
- Root-mean-square fluctuation (RMSF): Quantifies per-residue flexibility; binding pocket residues often show reduced fluctuation upon ligand binding [5]
- Hydrogen bond occupancy: Percentage of simulation time during which specific hydrogen bonds are maintained; high occupancy (>70%) indicates stable interactions [5]
- Binding free energy: MM-GBSA calculations provide quantitative estimates of binding affinity; potent STAT3 inhibitors typically show values ranging from -40 to -60 kcal/mol [5]
In studies of natural product inhibitors, lead compounds maintained stable binding poses throughout 100 ns simulations, with key hydrogen bonds to residues such as Arg609, Glu594, and Ser611 showing high occupancy (>80%) [5]. These quantitative metrics provide crucial validation of binding stability beyond initial docking scores.
Correlation with Experimental Data
The predictive power of MD simulations is greatly enhanced when correlated with experimental data. For SH2 domains, binding free energy models trained on high-throughput experimental data can achieve remarkable accuracy in predicting affinities for unseen peptide sequences [18]. One study using the ProBound statistical learning method achieved strong correlation (r² = 0.81) between predicted and experimental binding free energy parameters across different library designs [18]. This integration of computational and experimental approaches provides a robust framework for understanding SH2 domain specificity and designing targeted inhibitors.
Table 3: Key Residues in STAT3 SH2 Domain Binding Pocket
| Residue | Location | Role in Ligand Binding |
|---|---|---|
| Arg609 | βB strand | Forms critical salt bridge with phosphotyrosine [5] |
| Glu594 | αA helix | Participates in hydrogen bonding network [5] |
| Lys591 | αA helix | Contributes to electrostatic interactions [5] |
| Ser611 | BC loop | Forms hydrogen bonds with peptide backbone [5] |
| Ser636 | βD strand | Participates in sidechain recognition [5] |
| Tyr657 | EF loop | Contributes to hydrophobic interactions [5] |
| Gln644 | αB helix | Mediates specific sidechain recognition [5] |
Molecular dynamics simulations have revolutionized our understanding of STAT SH2 domain flexibility, providing insights that are transforming drug discovery approaches. As simulation methodologies continue to advance, several promising directions are emerging. The integration of machine learning with MD simulations shows particular promise, with sequence-to-affinity models like ProBound achieving impressive predictive accuracy for SH2 domain binding specificities [18]. Additionally, the recognition that SH2 domains can participate in liquid-liquid phase separation (LLPS) through multivalent interactions opens new avenues for therapeutic intervention [9].
The emerging understanding of non-canonical SH2 domain functions, including interactions with membrane lipids and roles in condensate formation, suggests that future MD studies should incorporate more complex biological environments [9]. Simulations that model SH2 domains in membrane-proximal contexts or within phase-separated condensates may reveal allosteric mechanisms and regulatory principles that could be exploited for more selective inhibition.
In conclusion, molecular dynamics simulations spanning nanosecond-to-microsecond timescales have provided unprecedented insights into the flexibility and function of STAT SH2 domains. By capturing the dynamic nature of these domains, MD simulations have enabled more effective virtual screening strategies, identified novel inhibitor candidates, and revealed fundamental mechanisms of SH2 domain function. As computational power continues to grow and methodologies refine, MD simulations will play an increasingly central role in targeting STAT3 and other challenging drug targets, ultimately accelerating the development of novel therapeutic agents for cancer and other diseases.
In the realm of structure-based drug discovery, the inherent flexibility of protein targets presents a formidable challenge. Conventional virtual screening often relies on static crystal structures, which may not accurately represent the dynamic conformational states that proteins adopt in solution. This limitation is particularly acute when targeting protein-protein interactions mediated by modular domains such as the STAT SH2 domain, where conformational flexibility is essential for function. The induced-active site strategy represents a paradigm shift that addresses this fundamental limitation by integrating molecular dynamics (MD) simulations to capture the dynamic behavior of therapeutic targets before screening compound libraries.
The STAT (Signal Transducers and Activators of Transcription) family of proteins, particularly STAT3, plays pivotal roles in cellular signaling pathways governing proliferation, survival, and differentiation. The SH2 (Src Homology 2) domain of STAT3 is especially critical for its function, facilitating recruitment to phosphorylated receptor complexes and mediating STAT3 dimerization through reciprocal phosphotyrosine-pTyr705-SH2 domain interactions [5] [21]. This dimerization is essential for STAT3 nuclear translocation and DNA binding, making the SH2 domain a highly attractive target for therapeutic intervention in cancers and inflammatory diseases characterized by constitutive STAT3 activation [4]. However, the SH2 domain exhibits considerable structural flexibility, with its phosphopeptide binding region resolved to only ~20 Å in crystal structures due to conformational dynamics [4]. This flexibility complicates drug discovery efforts, as static structures may not adequately represent the spectrum of conformations available for ligand binding.
The Induced-Active Site Methodology: A Technical Framework
The induced-active site strategy employs molecular dynamics simulations to generate a more physiologically relevant representation of the target's binding site. This approach recognizes that proteins are dynamic entities whose structural plasticity can significantly impact small molecule binding. The methodology involves a sequential process that transforms a static crystal structure into an ensemble of conformations for improved virtual screening.
Workflow Implementation
Table 1: Key Stages in the Induced-Active Site Strategy Implementation
| Stage | Process Description | Key Parameters | Primary Outcome |
|---|---|---|---|
| 1. System Preparation | Structure preparation of target protein complexed with high-affinity ligand | Selection of appropriate force field; solvation; energy minimization | Stable starting structure for MD simulation |
| 2. MD Simulation | Production run capturing thermodynamic fluctuations of the complex | Simulation time (ns); temperature (K); pressure (bar) | Trajectory file capturing temporal structural evolution |
| 3. Conformational Averaging | Extraction of representative structure from stable simulation phase | RMSD stabilization criteria; time frame selection (e.g., final 2ns) | "Averaged" structure reflecting induced-active site conformation |
| 4. Structure Optimization | Energy minimization of averaged structure | Implicit solvent model; convergence criteria | Refined receptor model for virtual screening |
| 5. Virtual Screening & Validation | Screening of compound libraries against induced-active site model | Docking algorithms; binding affinity scoring; interaction analysis | Identification of hit compounds with predicted bioactivity |
The foundational step in this methodology involves creating a dynamic model of the SH2 domain in complex with a known high-affinity ligand. In the case of STAT3 SH2 domain screening, researchers employed the peptidomimetic inhibitor CJ-887 (with a Kᵢ value of 15 nM) as the structuring ligand during MD simulations [4]. These simulations were conducted using the AMBER force field, with an explicit solvent model to better mimic physiological conditions. The production simulation typically extends for 10-20 nanoseconds, allowing adequate sampling of the conformational space accessible to the SH2 domain.
A critical innovation in this approach is the generation of an averaged structure derived from the MD trajectory, particularly from the period when the root mean square deviation (RMSD) has stabilized, indicating equilibrium conditions [4] [28]. This averaged structure is not simply a mathematical abstraction but represents a conformational state that has been "induced" through interaction with a binding partner and optimized through simulated thermodynamic sampling. The resulting model typically reveals subtle but critical rearrangements in side chain orientations and backbone adjustments that create potentially more druggable binding pockets compared to the static crystal structure.
Comparative Advantage Over Conventional Approaches
Traditional structure-based virtual screening typically relies on a single crystal structure as the receptor model, which represents just one snapshot from the ensemble of conformations the protein samples in solution. This static approach may fail to identify compounds that require specific induced conformations for binding, particularly for highly flexible domains like SH2. The induced-active site strategy addresses this fundamental limitation by capturing protein flexibility before the screening process begins.
This methodology proved particularly valuable for STAT3 inhibitor discovery, where previous screening efforts had identified small molecules with favorable drug-like properties but weak binding affinities, potentially due to the high flexibility of the target SH2 domain [4]. By using an MD-derived averaged structure that better represents the solution conformation when bound to a high-affinity ligand, researchers identified novel STAT3 inhibitors that interacted directly with key residues (R609 and S613) in the pY+0 binding pocket [4]. Notably, the hits identified through this approach were uncharged compounds with favorable drug-like properties, unlike most previous small-molecule STAT3 inhibitors that contained negatively-charged moieties to mimic phosphotyrosine [28].
Research Reagent Solutions for Implementation
Successful implementation of the induced-active site strategy requires specialized computational tools and biological reagents. The following table summarizes key resources employed in STAT3 SH2 domain screening.
Table 2: Essential Research Reagents and Computational Tools for Induced-Active Site Screening
| Category | Specific Resource | Application Purpose | Implementation Example |
|---|---|---|---|
| Target Structures | STAT3 SH2 domain crystal structures (e.g., 6NJS) | Provides initial coordinates for MD simulations | 6NJS selected for better resolution (2.70 Å) and unmutated SH2 domain [5] |
| Reference Ligands | High-affinity peptidomimetics (e.g., CJ-887) | Serves as structuring agent during MD simulations | CJ-887 (Kᵢ = 15 nM) used to induce biologically relevant conformations [4] |
| MD Software | AMBER, GROMACS, Desmond, YASARA | Performs molecular dynamics simulations | AMBER14 force field used for STAT3 SH2 simulations [4] |
| Docking Platforms | AutoDock Vina, GOLD, GLIDE, Schrödinger Suite | Conducts virtual ligand screening | SPECS database (110,000 compounds) screened against induced-active site [4] |
| Analysis Tools | PCA, FEL, MM/PBSA, MM/GBSA | Analyzes trajectories and calculates binding energies | MM/PBSA calculations validate binding affinities of hits [29] [30] |
Beyond these specialized resources, successful implementation requires access to high-performance computing infrastructure. The MD simulations central to this approach are computationally intensive, often requiring access to computing clusters or cloud-based resources. For reference, the simulation of the STAT3 SH2 domain in complex with CJ-887 utilized the BlueBioU high-performance computing resources at Rice University [4]. The emergence of large-scale quantum chemical datasets like Meta's Open Molecules 2025 (OMol25), which contains over 100 million molecular calculations, now provides unprecedented training data for refining neural network potentials that could accelerate such simulations [31] [32].
Experimental Protocol: STAT3 SH2 Domain Case Study
System Preparation and MD Simulation
The initial step involves preparing the protein-ligand complex for molecular dynamics simulation. For the STAT3 SH2 domain study, researchers began with the following protocol:
Structure Preparation: The STAT3 SH2 domain structure was obtained from the Protein Data Bank (preferably 6NJS for its better resolution and unmutated SH2 domain) [5]. The structure was processed using protein preparation tools to add hydrogen atoms, fill missing side chains, assign bond orders, and optimize hydrogen bonding networks.
Ligand Docking: The peptidomimetic inhibitor CJ-887 was docked into the SH2 domain binding site to establish the initial complex structure. The docking validation confirmed similar binding orientation to the STAT3 pY705 peptide motif, with critical interactions preserved in the pY+0 binding pocket [4].
Molecular Dynamics Simulation: The complex was solvated in an explicit water model, neutralized with appropriate ions, and energy-minimized before production dynamics. The simulation was conducted using the AMBER force field with the following parameters: simulation time of 10-20 nanoseconds, constant temperature (310 K), and constant pressure (1 bar) [4]. Particle Mesh Ewald method was employed for long-range electrostatic interactions, with a 2 femtosecond time step.
Trajectory Analysis and Model Generation
Following the MD simulation, the trajectory was analyzed to identify a stable simulation period and generate the induced-active site model:
Stability Assessment: The root mean square deviation (RMSD) of the protein backbone was calculated throughout the trajectory to identify when the system reached equilibrium. The final 2 nanoseconds of stable trajectory were typically selected for analysis [4].
Structure Averaging: An averaged structure was calculated from the stable simulation period, representing the "induced-active site" conformation. This structure was subsequently energy-minimized using implicit solvent models to remove any structural clashes introduced during the averaging process.
Binding Pocket Analysis: The induced-active site was compared with the original crystal structure to identify conformational changes, particularly in key residues like R609 and S613 in the pY+0 binding pocket of STAT3 [4].
Virtual Screening and Experimental Validation
The optimized induced-active site model served as the receptor for structure-based virtual screening:
Compound Library Screening: The SPECS database of 110,000 compounds was screened against the induced-active site model using docking software. The top 30% of hits were subjected to re-docking and re-scoring to improve ranking accuracy [4].
Hit Selection Criteria: Compounds were prioritized based on docking scores, binding mode analysis (particularly direct interactions with R609 and S613), and drug-like properties according to Lipinski's rule of five [4].
Experimental Validation: The top hits were tested for STAT3 inhibitory activity in cellular assays, including inhibition of cytokine-induced STAT3 tyrosine phosphorylation (pY-STAT3) and STAT3 DNA-binding activity [4]. The most promising compounds showed activity in the low micromolar range (2.7-34.5 µM) with favorable molecular properties for further optimization.
Advanced Applications and Future Directions
The induced-active site strategy represents a significant advancement in targeting challenging protein interfaces, but its implementation continues to evolve with emerging computational technologies. Recent developments in machine learning potentials trained on massive quantum chemical datasets promise to further enhance the accuracy and accessibility of MD simulations for drug discovery.
The release of resources like Meta's Open Molecules 2025 (OMol25) dataset, containing over 100 million molecular calculations at the ωB97M-V/def2-TZVPD level of theory, provides unprecedented training data for neural network potentials [31] [32]. These potentials can predict molecular energies and forces with DFT-level accuracy but at a fraction of the computational cost, potentially making long-timescale MD simulations more accessible for routine drug discovery applications. The Universal Models for Atoms (UMA) architecture, which unifies OMol25 with other datasets through a Mixture of Linear Experts approach, demonstrates how knowledge transfer across diverse chemical spaces can improve model performance [31].
Future applications of the induced-active site strategy will likely incorporate enhanced sampling techniques to more efficiently explore conformational space and identify rare but functionally relevant states. Additionally, the integration of machine learning approaches with physical simulations holds promise for accelerating both the MD simulations themselves and the subsequent virtual screening steps. As these technologies mature, the induced-active site strategy may become a standard approach for targeting not only SH2 domains but other challenging protein classes characterized by significant conformational flexibility, such as GPCRs, ion channels, and other modular interaction domains.
The Src Homology 2 (SH2) domain is a critical protein interaction module found in approximately 110 human proteins, including the Signal Transducer and Activator of Transcription 3 (STAT3) transcription factor [1] [9]. This domain specifically recognizes and binds to phosphorylated tyrosine residues, serving as a fundamental mechanism for signal transduction in eukaryotic cells [21]. In STAT3 signaling, the SH2 domain mediates the dimerization process essential for its activation and nuclear translocation, which promotes the expression of genes involved in cell proliferation, survival, and immune evasion [5] [33]. Dysregulated STAT3 activation is observed in numerous cancers, making its SH2 domain an attractive therapeutic target for cancer therapy [5] [34].
The discovery of inhibitors targeting protein-protein interactions like STAT3 dimerization presents considerable challenges due to the extensive, relatively shallow surface areas involved [21]. Computational approaches have emerged as powerful tools to address these challenges, enabling the efficient screening of vast chemical libraries to identify potential therapeutic candidates [5]. Natural products are particularly promising sources for drug discovery due to their inherent structural diversity, biological relevance, and favorable pharmacokinetic profiles compared to synthetic compounds [5]. Historical data indicates that approximately 40% of FDA-approved drugs are derived from natural sources, highlighting their therapeutic value [5].
This case study examines the application of computational methods for identifying natural compound inhibitors of the STAT3-SH2 domain from large databases, framed within broader research on the molecular dynamics and flexibility of STAT SH2 domains.
STAT3 Signaling and SH2 Domain Structure
STAT3 Activation Pathway
STAT3 activation is initiated by various extracellular signals, including cytokines and growth factors. This activation triggers a phosphorylation cascade that ultimately leads to STAT3 dimerization and nuclear translocation. The accompanying diagram illustrates this signaling pathway and the critical role of the SH2 domain in facilitating protein-protein interactions that drive oncogenic processes.
SH2 Domain Architecture and Binding Pockets
The SH2 domain adopts a conserved three-dimensional structure characterized by a central anti-parallel β-sheet flanked by two α-helices, forming an αββββα motif [5] [9]. Despite relatively low sequence identity among some family members (as little as ~15%), all SH2 domains assume nearly identical folds, suggesting these folds have evolved almost exclusively to bind phosphotyrosine-containing motifs [9].
Structurally, SH2 domains can be divided into two major subgroups: the SRC type and STAT type. STAT-type SH2 domains are distinct in that they lack the βE and βF strands as well as the C-terminal adjoining loop, with the αB helix split into two helices [9]. This structural disparity likely represents an adaptation that facilitates STAT dimerization, reflecting the ancestral function of SH2 domain-containing proteins that predate animal multicellularity [9].
The phosphotyrosine (pY) binding pocket of the STAT3 SH2 domain is divided into three principal sub-pockets that serve as key binding sites for inhibitors:
- pY+0 pocket: Binds directly to phosphotyrosine 705 (pY705) and contains polar residues responsible for hydrogen bonding and electrostatic interactions [5] [35]
- pY+1 pocket: Interacts with leucine 706 (L706) and is formed predominantly by hydrophobic residues [5]
- pY-X pocket: A hydrophobic side pocket that provides additional binding specificity [33] [35]
These sub-pockets create an extended binding surface that recognizes the phosphotyrosine motif and facilitates STAT3 dimerization. Key residues involved in binding include Arg609, Glu594, Lys591, Ser636, Ser611, Val637, Tyr657, Gln644, Thr640, Glu638, and Trp623 [5]. The high conservation of the pY+0 binding pocket across STAT family members presents challenges for developing specific inhibitors that avoid cross-reactivity [33].
Computational Workflow for Inhibitor Identification
The identification of natural compound inhibitors from large databases follows a multi-stage computational workflow that progressively filters candidates based on binding affinity, pharmacological properties, and complex stability. The following diagram illustrates this sequential screening process:
Database Preparation and Virtual Screening
The initial stage involves curating a comprehensive library of natural compounds for screening:
Database Source and Preparation:
- Source: 182,455 natural compounds retrieved from the ZINC15 database under "now availability" criteria [5]
- Preparation: Compound structures were processed using LigPrep (Schrödinger Suite) to generate optimized 3D structures with correct ionization states at physiological pH (7.4 ± 0.5) [5]
- Optimization: Molecular structures were further refined using the OPLS3e force field to ensure proper energetics and stereochemistry [5]
Virtual Screening Protocol: Virtual screening employed a multi-tiered docking approach using the GLIDE module to progressively identify high-affinity binders:
- High-Throughput Virtual Screening (HTVS): Initial rapid screening of all 182,455 prepared ligands to identify promising candidates
- Standard Precision (SP) Docking: Refined docking of the top 55,872 compounds (approximately 30%) identified through HTVS
- Extra Precision (XP) Docking: High-accuracy docking of the most promising compounds (cut-off at -6.5 kcal/mol) from the SP stage [5]
Receptor Grid Preparation: The crystal structure of STAT3 (PDB ID: 6NJS) was selected for docking studies based on its superior resolution (2.70 Å), absence of mutations in the SH2 domain, and fewer sequence gaps compared to alternative structures [5]. The receptor grid was generated centered on the coordinates X:13.22, Y:56.39, Z:0.27 with a box size of 20 Å, encompassing the key sub-pockets of the SH2 domain [5].
Binding Affinity and Free Energy Calculations
Compounds exhibiting favorable docking scores advanced to more rigorous binding affinity assessment:
MM-GBSA Calculations: The Molecular Mechanics Generalized Born Surface Area (MM-GBSA) method was employed to calculate binding free energies using the Prime module (Schrödinger Suite) [5]. This approach combines molecular mechanics calculations with implicit solvation models to provide more accurate binding affinity estimates than docking scores alone.
The binding free energy (ΔGBinding) was calculated using the equation: ΔGBinding = ΔGComplex - (ΔGReceptor + ΔGLigand)
where ΔGComplex, ΔGReceptor, and ΔGLigand represent the free energies of the protein-ligand complex, free receptor, and unbound ligand, respectively [5]. More negative ΔGBinding values indicate stronger binding potential.
Pharmacokinetic Property Prediction: The QikProp tool was utilized to assess drug-like properties and pharmacokinetic profiles of candidate compounds, applying criteria such as Lipinski's Rule of Five to prioritize molecules with higher probability of clinical success [5].
Molecular Dynamics Simulations
Molecular dynamics (MD) simulations provide critical insights into the stability and conformational dynamics of protein-ligand complexes under conditions mimicking the physiological environment:
Simulation Parameters:
- Software: Desmond MD system (Schrödinger Suite) or GROMACS [5] [36]
- Force Field: OPLS3e or GAFF for small molecules [5] [34]
- Solvation Model: SPC water model in a cubic box [34]
- Neutralization: Appropriate ions added to neutralize system charge [34]
- Simulation Time: 100 ns production run following system equilibration [36]
Trajectory Analysis: The stability of protein-ligand complexes was assessed through multiple analytical approaches:
- Root Mean Square Deviation (RMSD): Measures structural stability over time, with values below 0.3 nm indicating stable complexes [35]
- Root Mean Square Fluctuation (RMSF): Evaluates residual flexibility and identifies regions of structural instability
- Hydrogen Bond Analysis: Quantifies persistent intermolecular interactions critical for binding
- Binding Mode Evolution: Assesses whether the ligand maintains its initial binding pose throughout simulation
Advanced Analyses:
- WaterMap Analysis: Identifies the thermodynamic properties of hydration sites within the binding pocket [5]
- Principal Component Analysis: Identifies essential collective motions of the protein-ligand complex
- Dynamic Cross-Correlation Analysis: Maps correlated motions between different protein regions
Specificity Assessment and Network Pharmacology
Given the high conservation of SH2 domains across STAT family members, specificity assessment is crucial for inhibitor development:
Cross-Binding Evaluation: Candidate inhibitors were docked against structurally related SH2 domains (particularly STAT1) to identify compounds with selective binding profiles [33]. This evaluation helps minimize off-target effects that could compromise therapeutic utility.
Network Pharmacology: Mapping compound-target interactions within broader biological networks helps elucidate multi-target potential and identify potential synergistic effects or unintended pathway modulations [5]. This systems biology approach provides a more comprehensive understanding of a compound's pharmacological profile beyond single-target activity.
Results and Validation
Identified Natural Compound Inhibitors
The computational screening pipeline identified several natural compounds with promising potential as STAT3-SH2 domain inhibitors:
Table 1: Promising Natural Compound Inhibitors of STAT3-SH2 Domain
| Compound ID | Source | Docking Score (kcal/mol) | Key Binding Residues | Cellular IC50 | Reference |
|---|---|---|---|---|---|
| ZINC67910988 | Natural Product Database | -9.8 | Lys591, Glu594, Arg609, Ser611 | Under investigation | [5] |
| ZINC255200449 | Natural Product Database | -9.2 | Lys591, Glu594, Ser636 | Under investigation | [5] |
| ZINC299817570 | Natural Product Database | -8.9 | Lys591, Gln644, Thr640 | Under investigation | [5] |
| ZINC31167114 | Natural Product Database | -8.7 | Glu638, Trp623, Tyr657 | Under investigation | [5] |
| (-)-Epigallocatechin gallate | JAK/STAT Library | -10.2 | pY+0 and pY+1 pocket residues | Low micromolar range | [36] |
| Kaempferol-3-O-rutinoside | JAK/STAT Library | -9.8 | pY+0 and pY+1 pocket residues | Low micromolar range | [36] |
| Saikosaponin D | JAK/STAT Library | -9.5 | pY+0 and pY-X pocket residues | Low micromolar range | [36] |
| PMM-172 | Shikonin derivative | -8.9 | Lys591, Glu594, Ile634, Arg595 | 1.98 ± 0.49 μM (MDA-MB-231) | [35] |
Molecular Dynamics Validation
Molecular dynamics simulations provided critical validation of compound stability and binding mechanisms:
Table 2: Molecular Dynamics Analysis of Top Candidates
| Compound ID | RMSD (nm) | Hydrogen Bonds | Key Interactions | Simulation Time |
|---|---|---|---|---|
| ZINC67910988 | < 0.2 | 4-6 persistent | Stable in pY+0 and pY+1 pockets | 100 ns |
| PMM-172 | < 0.15 | 3-5 persistent | Maintained hydrogen bonds with Lys591, Glu594, Arg595 | 100 ns |
| (-)-Epigallocatechin gallate | < 0.25 | 5-7 persistent | Stable binding in multiple subpockets | 100 ns |
| Stattic (control) | 0.2-0.35 | 2-4 intermittent | Moderate stability with some positional shifts | 100 ns |
ZINC67910988 demonstrated superior stability in molecular dynamics simulations, maintaining its binding pose with minimal fluctuation throughout the 100 ns simulation period [5]. PMM-172, a shikonin derivative, also showed exceptional stability, rapidly reaching equilibrium and maintaining it for over 3 ns with minimal structural deviation [35]. This compound formed additional hydrogen bonds with residue Arg595 compared to its parent scaffold, explaining its improved binding affinity [35].
Experimental Validation
Promising computational hits typically advance to experimental validation to confirm biological activity:
Cellular Efficacy Assessment:
- Anti-proliferative Assays: Candidate compounds are tested against cancer cell lines with constitutive STAT3 activation (e.g., MDA-MB-231 breast cancer cells) [35]
- Apoptosis Induction: Flow cytometry with Annexin V/PI staining quantifies programmed cell death [35]
- STAT3 Phosphorylation Inhibition: Western blotting detects reduced Tyr705 phosphorylation following compound treatment [34]
- STAT3 Transcriptional Activity: Luciferase reporter assays measure inhibition of STAT3-dependent transcription [35]
PMM-172 demonstrated potent anti-proliferative activity against triple-negative breast cancer cells (MDA-MB-231) with an IC50 of 1.98 ± 0.49 μM, outperforming the natural compound shikonin (IC50 = 2.88 ± 0.25 μM) from which it was derived [35]. This compound also induced dose-dependent apoptosis, with 62.74% of cells undergoing apoptosis at 8 μM concentration, and effectively inhibited STAT3 nuclear localization and downstream target gene expression [35].
The Scientist's Toolkit
Successful implementation of this case study requires specialized software tools and databases:
Table 3: Essential Research Tools for Computational Inhibitor Screening
| Tool/Database | Type | Primary Function | Application in Workflow |
|---|---|---|---|
| ZINC15 | Chemical Database | Source of commercially available natural compounds | Initial compound library generation [5] |
| Schrödinger Suite | Software Platform | Integrated computational drug discovery platform | Protein preparation, docking, MD simulations [5] |
| GROMACS | Software Tool | Molecular dynamics simulation package | MD simulations and trajectory analysis [36] |
| Wordom | Software Tool | Molecular simulation analysis | Analysis of conformational ensembles from MD [37] |
| RCSB PDB | Protein Database | Source of 3D protein structures | Retrieval of STAT3 crystal structure (6NJS) [34] |
| QikProp | Software Tool | ADME prediction | Pharmacokinetic property assessment [5] |
| GLIDE | Software Tool | Molecular docking | HTVS, SP, and XP docking simulations [5] |
| Desmond | Software Tool | Molecular dynamics | MD simulations and stability analysis [5] |
This case study demonstrates a robust computational framework for identifying natural compound inhibitors of the STAT3-SH2 domain from large databases. The multi-stage screening approach—progressing from virtual screening to molecular dynamics simulations—effectively prioritizes candidates with favorable binding characteristics, stability, and drug-like properties. The identification of compounds such as ZINC67910988 and PMM-172 highlights the potential of natural products as starting points for developing targeted cancer therapeutics.
The integration of molecular dynamics simulations provides critical insights into SH2 domain flexibility and inhibitor complex stability, enabling more accurate prediction of biological activity. These computational approaches significantly accelerate the early drug discovery process by prioritizing the most promising candidates for experimental validation, ultimately reducing the time and resources required for therapeutic development.
Future directions in this field will likely involve more sophisticated simulations capturing longer timescales, incorporation of enhanced sampling techniques to explore rare conformational events, and integration of machine learning approaches to further improve screening efficiency. As our understanding of SH2 domain dynamics advances, so too will our ability to design selective inhibitors that disrupt pathological STAT3 signaling while minimizing off-target effects.
Src homology 2 (SH2) domains are approximately 100-amino-acid protein modules that specifically recognize and bind phosphorylated tyrosine (pY) motifs, serving as crucial components in intracellular signal transduction networks [16] [9]. These domains are found in approximately 110 human proteins with diverse cellular functions, including enzymes, adaptor proteins, transcription factors, and cytoskeletal proteins [16] [38]. The canonical function of SH2 domains involves facilitating protein-protein interactions by recruiting specific binding partners to activated receptor tyrosine kinases, thereby propagating downstream signaling cascades.
Traditional structural biology approaches, particularly X-ray crystallography, have provided foundational insights into SH2 domain architecture. These domains consistently adopt a characteristic "sandwich" fold consisting of a central three-stranded antiparallel beta-sheet flanked by two alpha helices (αA-βB-βC-βD-αB) [16] [9]. The N-terminal region contains a deeply conserved pY-binding pocket featuring an invariant arginine residue (βB5) that forms a salt bridge with the phosphate moiety of phosphorylated tyrosine residues [9]. Despite these structural conserved features, SH2 domains exhibit remarkable specificity in recognizing distinct pY-containing motifs, primarily determined by residues C-terminal to the phosphotyrosine.
While static structures have been invaluable for understanding basic SH2 domain architecture, they provide limited insights into the conformational dynamics and allosteric regulation that govern SH2 function in physiological contexts. This limitation is particularly relevant for STAT (Signal Transducer and Activator of Transcription) proteins, whose SH2 domains mediate critical dimerization events essential for their function as transcription factors [39]. The emergence of molecular dynamics (MD) simulations and complementary computational approaches has enabled researchers to transition from analyzing static structures to characterizing dynamic ensembles, revealing previously unappreciated allosteric networks and hidden conformational states with significant implications for therapeutic development.
Structural Fundamentals of SH2 Domains
Conserved Architecture with Functional Diversification
All SH2 domains share a structurally conserved core fold despite significant sequence variation, with some family members sharing as little as 15% pairwise sequence identity [16] [9]. This structural conservation suggests the fold has been optimized specifically for phosphotyrosine recognition throughout evolution. The central β-sheet forms the structural backbone, while the surrounding α-helices and connecting loops contribute to binding specificity and regulatory potential.
Table 1: SH2 Domain Structural Elements and Their Functional Roles
| Structural Element | Description | Functional Role |
|---|---|---|
| βB strand | Central beta strand | Contains invariant arginine (βB5) for pY binding |
| FLVR motif | Highly conserved sequence motif | Forms phosphate-binding pocket |
| pY pocket | Deep pocket near N-terminus | Binds phosphotyrosine moiety |
| Specificity pocket | Adjacent to pY pocket | Recognizes residues C-terminal to pY |
| EF loop | Connects βE and βF strands | Determines ligand access and selectivity |
| BG loop | Connects αB helix and βG strand | Regulates binding specificity |
| CD loop | Variable length loop | Contributes to functional diversity |
STAT-type SH2 domains represent a structurally distinct subgroup characterized by the absence of βE and βF strands and a split αB helix [9]. This structural adaptation likely facilitates the dimerization process essential for STAT-mediated transcriptional activation. The specialized architecture of STAT SH2 domains enables reciprocal phosphotyrosine-SH2 interactions that stabilize active dimers, a mechanism crucial for proper STAT function in JAK-STAT signaling pathways.
Beyond Phosphopeptide Binding: Emerging Regulatory Mechanisms
Recent research has revealed that SH2 domains participate in more complex regulatory mechanisms than previously appreciated:
Lipid Binding: Approximately 75% of SH2 domains interact with membrane lipids, particularly phosphatidylinositol-4,5-bisphosphate (PIP₂) and phosphatidylinositol-3,4,5-trisphosphate (PIP₃) [16] [9]. These interactions often involve cationic regions near the pY-binding pocket flanked by aromatic or hydrophobic residues, enabling membrane recruitment and modulation of enzymatic activity.
Liquid-Liquid Phase Separation (LLPS): SH2 domains contribute to the formation of biomolecular condensates through multivalent interactions [16]. For example, interactions among GRB2, Gads, and LAT receptors drive LLPS formation that enhances T-cell receptor signaling [16].
Allosteric Regulation: SH2 domains can transmit conformational changes across large distances, as demonstrated in STAT3, where perturbations in the coiled-coil domain (CCD) allosterically regulate SH2 domain conformation and function [39].
Computational Methodologies for Mapping Dynamic Ensembles
Molecular Dynamics Simulations: Technical Framework
Molecular dynamics (MD) simulations provide atomic-resolution insights into protein dynamics by numerically solving Newton's equations of motion for all atoms in a system. The following protocol outlines a standardized approach for investigating SH2 domain dynamics:
Table 2: Molecular Dynamics Simulation Protocol for SH2 Domain Analysis
| Step | Parameter | Specification | Purpose |
|---|---|---|---|
| 1. System Preparation | Protein Structure | PDB ID (e.g., 6CRF for SHP2) [40] | Initial coordinates |
| Solvation | TIP3P water model, 10-15 Å padding | Hydration environment | |
| Neutralization | Ionic concentration (e.g., 150 mM NaCl) | Physiological conditions | |
| 2. Force Field Selection | Protein | CHARMM36/AMBER ff19SB | Atomic interactions |
| Lipids | SLIPIDS/CHARMM36 | Membrane simulations | |
| 3. Simulation Parameters | Integration | 2-fs time step | Numerical stability |
| Temperature | 310 K (Nose-Hoover thermostat) | Physiological temperature | |
| Pressure | 1 bar (Parrinello-Rahman barostat) | Isotropic-isothermal conditions | |
| Non-bonded | Particle Mesh Ewald (PME) | Long-range electrostatics | |
| 4. Production Simulation | Duration | 100 ns - 1 μs (equilibrium MD) [41] | Conformational sampling |
| Replicates | 3+ independent trajectories | Statistical significance | |
| 5. Enhanced Sampling | Method | Meta-dynamics [41]/Replica Exchange [40] | Free energy landscape |
Enhanced Sampling Techniques
Conventional MD simulations may insufficiently sample rare conformational transitions relevant to allosteric regulation. Enhanced sampling methods address this limitation:
Meta-dynamics Simulations: This approach accelerates sampling by adding history-dependent bias potentials that discourage revisiting previously explored configurations [41]. Applied to SHP2, meta-dynamics has revealed free energy landscapes underlying the transition between autoinhibited and active states, identifying metastable intermediate states not observed in crystal structures [41].
Replica Exchange MD (REMD): Also known as parallel tempering, REMD facilitates barrier crossing by simulating multiple copies of the system at different temperatures and periodically exchanging configurations between temperatures [40]. This approach has been used to demonstrate that the crystallographic active state of SHP2 is unstable in solution, revealing multiple interdomain arrangements that facilitate association with bisphosphorylated sequences [40].
Binding Free Energy Calculations
Quantifying interaction energetics is crucial for understanding allosteric regulation and inhibitor binding:
MM/GBSA Method: The Molecular Mechanics/Generalized Born Surface Area approach estimates binding free energies by combining molecular mechanics energy terms with continuum solvation models [41]. This method has been applied to characterize the interactions of 45 allosteric inhibitors with SHP2, revealing thermodynamic determinants of binding affinity [41].
Potential of Mean Force (PMF): PMF calculations provide absolute binding free energies by sampling along a reaction coordinate, offering insights into specificity determinants of SH2 domain-phosphopeptide interactions [42].
Case Study: Allosteric Regulation of STAT3 SH2 Domain
Experimental Evidence for Allosteric Communication
STAT3 represents a paradigm for allosteric regulation in SH2 domain-containing proteins. The protein consists of six domains: N-terminal domain (NTD), coiled-coil domain (CCD), DNA-binding domain (DBD), linker domain (LD), SH2 domain, and transactivation domain (TAD) [39]. Several lines of evidence demonstrate allosteric communication between the CCD and SH2 domain:
- The D170A mutation in CCD diminishes both phosphopeptide binding and tyrosine phosphorylation without directly affecting the SH2 domain [39].
- Small molecule inhibitors (e.g., MM-206, K116) binding to CCD allosterically inhibit SH2 domain function and STAT3 phosphorylation [39].
- A small polypeptide (MS3-6) binding to CCD induces significant helical tilts that diminish DNA binding and nuclear translocation [39].
Molecular Dynamics Reveal the Allosteric Pathway
MD simulations of wild-type STAT3 and the D170A variant have elucidated the structural basis for allosteric communication between CCD and SH2 domains [39]. The analysis reveals:
- Rigid Core Transmission: Perturbations in CCD are transmitted through a rigid backbone core connecting CCD and SH2 via the linker domain (LD).
- Specific Residue Networks: The allosteric pathway involves a network of short-range interactions that propagate conformational changes from CCD to SH2.
- SH2 Conformational Changes: The D170A variant exhibits distinctive conformational changes in the SH2 domain, particularly in residues comprising the pY pocket (R609, K591, S636, S611) and pY+3 pocket (V637, Y657, Q644, Y640, E638) [39].
Machine Learning for High-Dimensional Data Analysis
Interpretable Machine Learning in MD Analysis
The high-dimensional data generated by MD simulations presents analytical challenges that can be addressed through machine learning approaches:
Feature Extraction: Trajectory analysis data, ligand-receptor interaction fingerprints, and residue contact matrices serve as input features for machine learning models [41].
XGBoost with SHAP: The extreme gradient boosting (XGBoost) model combined with Shapley Additive Explanations (SHAP) provides an interpretable framework for identifying key structural features driving conformational dynamics [41]. This approach has been successfully applied to identify residues and interactions controlling SHP2 conformational changes and allosteric inhibitor activity [41].
Allosteric Pocket Prediction Workflow
The integration of MD simulations with machine learning enables robust prediction of allosteric pockets:
Experimental Validation and Research Applications
The Scientist's Toolkit: Essential Research Reagents
Table 3: Essential Research Reagents for SH2 Domain Allostery Studies
| Reagent/Category | Specification | Research Application |
|---|---|---|
| Structural Databases | SH2db [43], PDB, AlphaFold | Structural templates, sequence-structure analysis |
| MD Software | GROMACS, AMBER, NAMD, OpenMM | Molecular dynamics simulations |
| Enhanced Sampling | PLUMED, COLVAR | Meta-dynamics, replica exchange simulations |
| Analysis Tools | MDTraj, MDAnalysis, PyEMMA | Trajectory analysis, feature extraction |
| Machine Learning | XGBoost, SHAP, Scikit-learn | Predictive modeling, feature importance |
| SH2 Domain Library | Non-redundant human SH2 clones [38] | Functional screening, binding assays |
| Binding Assays | ITC, SPR, FP | Quantitative binding affinity measurement |
Therapeutic Applications and Drug Discovery
The identification and characterization of allosteric pockets in SH2 domains has significant therapeutic implications:
Allosteric Inhibitor Development: SHP2 allosteric inhibitors (e.g., SHP099, RMC-4630, TNO155) stabilize the autoinhibited conformation by binding at the interface of the C-SH2 and PTP domains, representing a promising therapeutic strategy for cancer treatment [41]. Eight allosteric SHP2 inhibitors have entered clinical trials for cancer therapy as of 2024 [41].
STAT3 Targeted Therapeutics: Allosteric modulation of STAT3 via its CCD domain offers an alternative to direct SH2 domain targeting, potentially overcoming specificity and pharmacologic efficacy challenges that have hampered clinical development [39].
Resistance Management: Allosteric inhibitors may offer advantages in addressing drug resistance issues, as allosteric sites are typically less conserved than orthosteric catalytic sites [41] [16].
The transition from analyzing static structures to characterizing dynamic ensembles has fundamentally advanced our understanding of SH2 domain function and allosteric regulation. Molecular dynamics simulations, enhanced sampling techniques, and interpretable machine learning collectively provide a powerful framework for predicting and validating allosteric pockets in SH2 domains and beyond. The integration of these computational approaches with experimental validation offers a robust strategy for identifying novel therapeutic targets and developing allosteric modulators with improved specificity and reduced potential for resistance.
Future advances in this field will likely involve longer timescale simulations enabled by computational hardware improvements, more sophisticated enhanced sampling algorithms, and the integration of multi-scale modeling approaches that bridge atomic-level simulations with cellular-scale signaling networks. As these methodologies continue to mature, they will increasingly guide rational drug design efforts targeting allosteric sites in challenging therapeutic targets like STAT SH2 domains.
Overcoming Challenges in Simulating and Targeting Flexible SH2 Domains
Addressing High Domain Mobility and Conformational Heterogeneity
High domain mobility and conformational heterogeneity are fundamental characteristics of proteins that govern their function, regulation, and interactions in cellular signaling pathways. Within the context of STAT (Signal Transducer and Activator of Transcription) proteins, the Src Homology 2 (SH2) domain exemplifies these dynamic properties, presenting both challenges and opportunities for research and therapeutic development. The SH2 domain, approximately 100 amino acids in length, is a modular protein interaction domain that specifically recognizes phosphotyrosine (pY) motifs, playing a crucial role in tyrosine kinase signaling networks [9]. Understanding the structural plasticity and dynamic behavior of STAT SH2 domains is essential for deciphering their biological functions and developing targeted therapeutic interventions for diseases such as cancer, where STAT signaling is frequently dysregulated.
This technical guide provides an in-depth examination of the core principles and methodologies for studying domain mobility and conformational heterogeneity in STAT SH2 domains. We explore the structural basis of SH2 domain flexibility, present experimental and computational approaches for characterizing dynamic behavior, and discuss implications for drug discovery efforts targeting these critical signaling domains.
Structural Fundamentals of SH2 Domains
Core Architecture and Flexibility
SH2 domains maintain a conserved structural fold despite significant sequence variation across different proteins. The canonical SH2 domain structure consists of a central three-stranded antiparallel beta-sheet flanked by two alpha helices, forming an αββα sandwich structure [9] [23]. This scaffold creates a specialized binding pocket for phosphorylated tyrosine recognition while maintaining inherent flexibility that enables functional diversity.
The N-terminal region of SH2 domains is highly conserved and contains a deep pocket within the βB strand that binds the phosphate moiety of phosphotyrosine. This pocket harbors an invariant arginine residue (at position βB5) that is part of the FLVR motif found in most SH2 domains and directly interacts with the phosphotyrosine through a salt bridge [9]. The C-terminal region displays greater variability and contains additional structural elements, including β-strands E, F, and G in many SH2 domains. The intervening loops between these structural elements, particularly the CD-loop, EF-loop, and BG-loop, vary in length and conformation across different SH2 domain families, contributing to ligand specificity and functional diversity [9].
Table 1: Key Structural Elements of SH2 Domains
| Structural Element | Description | Functional Role |
|---|---|---|
| Central β-sheet | Three antiparallel β-strands (βB-βD) | Structural scaffold forming binding surface |
| Flanking α-helices | Two α-helices (αA and αB) | Stabilize domain structure and contribute to binding surface |
| pY binding pocket | Deep pocket in βB strand containing FLVR motif | Recognizes and binds phosphotyrosine residues |
| BG loop | Loop between α-helix B and β-strand G | Contributes to conformational changes during activation |
| EF loop | Loop between β-strands E and F | Participates in phosphopeptide binding and specificity |
Molecular Basis of Conformational Heterogeneity
The conformational heterogeneity of SH2 domains arises from several structural features that enable dynamic behavior. Loop regions connecting secondary structural elements exhibit inherent flexibility, allowing adaptation to different binding partners. For example, in the Drk-SH2 domain (a GRB2 homologue), loops A, C, E, and F show considerable conformational variation compared to related structures, contributing to its dynamic behavior [44]. These flexible loops undergo structural rearrangements upon ligand binding, facilitating specific recognition of phosphopeptide motifs.
Allosteric networks within the SH2 domain structure enable communication between distant sites. Molecular dynamics simulations of various SH2-containing proteins reveal that conformational changes in one region can propagate throughout the domain structure. For instance, in SHP2, a protein tyrosine phosphatase containing two SH2 domains, the BG loop of the N-SH2 domain plays a previously underappreciated role in activation by mediating conformational changes that expose the binding site [40]. Similarly, studies on BTK (Bruton's tyrosine kinase) demonstrate conformational heterogeneity in its PHTH domain, which adopts a range of states arrayed around the autoinhibited SH3-SH2-kinase core [45] [46].
Experimental Methodologies for Characterizing Dynamics
Structural Biology Approaches
Cryo-Electron Microscopy (cryo-EM) has emerged as a powerful technique for visualizing conformational heterogeneity in multidomain proteins. Unlike X-ray crystallography, which often fails to resolve highly flexible regions, cryo-EM can capture multiple conformational states within a single sample. For full-length BTK, cryo-EM reconstructions provided the first view of the PHTH domain within the full-length protein, revealing that the globular PHTH domain adopts a range of states arrayed around the autoinhibited SH3-SH2-kinase core [45] [46]. This conformational heterogeneity had been refractory to crystallization attempts, with diffraction data showing only the structured core and no electron density for the flexible PHTH-PRR segment.
Solution NMR Spectroscopy offers unparalleled insights into protein dynamics at atomic resolution. NMR studies of the Drk-SH2 domain in complex with a phosphotyrosine-containing peptide from the Sevenless receptor revealed both the structure and dynamics of the domain [44]. The assignment of backbone and sidechain NMR resonances, combined with relaxation experiments, provided information on site-specific mobility and conformational exchange processes. Notably, the Drk-SH2 domain exhibited stability issues and concentration-dependent aggregation in the absence of binding partners, highlighting the intrinsic flexibility of this domain [44].
Table 2: Experimental Techniques for Studying SH2 Domain Dynamics
| Technique | Applications | Resolution | Time Scale | Limitations |
|---|---|---|---|---|
| Cryo-EM | Visualization of conformational states in full-length proteins | Near-atomic to intermediate | Static snapshots | Limited resolution for flexible regions |
| NMR Spectroscopy | Atomic-resolution dynamics, chemical environment, relaxation | Atomic | Picoseconds to seconds | Protein size limitations |
| HDX-MS | Protein dynamics, conformational changes, allostery | Peptide level | Milliseconds to hours | Indirect structural information |
| SAXS | Low-resolution shape, flexibility, oligomerization | Low resolution | Ensemble average | Modeling ambiguity |
Biophysical and Biochemical Assays
Hydrogen/Deuterium Exchange Mass Spectrometry (HDX-MS) provides information on protein dynamics by measuring the exchange rate of backbone amide hydrogens with solvent deuterium. This technique has been applied to study full-length BTK, revealing autoinhibitory interactions between the PHTH domain and the activation loop face of the BTK kinase domain that were not apparent in static structures [45] [46]. By comparing exchange patterns between different functional states, HDX-MS can map conformational changes and allosteric networks in SH2 domain-containing proteins.
High-Throughput Affinity Profiling enables comprehensive characterization of SH2 domain binding specificity and the energetic contributions of different peptide positions. Recent advances combine bacterial surface display of genetically-encoded peptide libraries with deep sequencing to quantify binding enrichment across thousands of candidate ligands [18]. This approach has been used to develop quantitative models of SH2 domain specificity, such as free energy matrices that predict binding affinity for any ligand sequence in the theoretical space covered by the library. These models reveal that SH2 domains exhibit distinct sequence preferences despite structural homology, reflecting functional specialization [18].
Computational and Modeling Approaches
Molecular Dynamics Simulations
Molecular Dynamics (MD) Simulations provide atomic-resolution insights into protein motions and conformational transitions. MD simulations have been extensively applied to study SH2 domain dynamics and allosteric regulation. For example, enhanced sampling simulations of SHP2 revealed that the crystallographic conformation of the active state is unstable in solution, with the protein populating multiple interdomain arrangements that facilitate association with bisphosphorylated sequences [40]. These simulations demonstrated that activation is coupled to conformational changes of the N-SH2 binding site, which becomes significantly more accessible in the active state.
Enhanced Sampling Techniques, such as meta-dynamics simulations, enable the exploration of conformational landscapes and free energy calculations. In studies of SHP2, meta-dynamics simulations provided insights into the free energy landscapes of apo and inhibitor-bound states, revealing stable, metastable, and transition states along the activation pathway [41]. These approaches have identified key structural features driving SHP2 conformational dynamics and regulating allosteric inhibitor activity, providing crucial insights for designing potent inhibitors and addressing drug resistance.
Table 3: Computational Methods for Studying SH2 Domain Dynamics
| Method | Application | Time Scale | Advantages | Requirements |
|---|---|---|---|---|
| Classical MD | Conformational sampling, loop dynamics | Nanoseconds to microseconds | Atomic resolution | High-performance computing |
| Enhanced Sampling (Meta-dynamics) | Free energy landscapes, rare events | Effectively extends to milliseconds | Accelerates barrier crossing | Careful parameter selection |
| MM/GBSA | Binding free energy calculations | End-point method | Computational efficiency | Ensemble of structures |
| Machine Learning (XGBoost) | Feature importance, conformational analysis | Trajectory analysis | Identifies key determinants | Large training datasets |
Integrative Modeling and Machine Learning
Interpretable Machine Learning approaches are increasingly applied to extract meaningful information from high-dimensional simulation data. In studies of SHP2, researchers employed extreme gradient boosting (XGBoost) with Shapley additive explanations (SHAP) to analyze molecular dynamics simulation trajectories and identify key residues and interactions controlling conformational changes [41]. This approach successfully handled complex protein structural dynamic information, reduced data dimensionality, and highlighted specific atoms or residues with significant impacts on protein conformation evolution.
Sequence-to-Affinity Models represent another application of computational approaches to understand SH2 domain function. The ProBound method uses statistical learning to build free-energy matrices from high-throughput protein-peptide binding data, enabling accurate prediction of binding affinity for any ligand sequence [18]. These models demonstrate superior robustness compared to traditional enrichment-based analyses and provide biophysically interpretable parameters (ΔΔG/RT) that are consistent across different library designs.
Case Studies: STAT3 and Related SH2 Domains
STAT3 SH2 Domain in Health and Disease
The STAT3 SH2 domain plays a critical role in STAT3 activation and dimerization, processes essential for its function as a transcription factor. STAT3 activation involves phosphorylation at tyrosine 705 (Y705), which promotes SH2 domain-mediated dimerization through reciprocal interactions between the phosphotyrosine of one STAT3 molecule and the SH2 domain of another [23]. This dimerization is essential for nuclear translocation and DNA binding, making the SH2 domain a compelling target for therapeutic intervention in cancer and other diseases characterized by aberrant STAT3 signaling.
The structural organization of the STAT3 SH2 domain follows the canonical SH2 fold but contains unique features that determine its specificity. The pY binding pocket is divided into three sub-pockets: the pY+X (hydrophobic side), pY+0 (binds to pY705), and pY+1 (binds to L706) pockets [23]. Key residues involved in binding include Arg609, Glu594, Lys591, Ser636, Ser611, Val637, Tyr657, Gln644, Thr640, Glu638, and Trp623. Mutations or disruptions in these residues can attenuate STAT3 signaling and activation, highlighting their functional importance.
Targeting STAT3 SH2 Domain for Therapeutic Intervention
Computational Screening approaches have been successfully applied to identify natural compounds targeting the SH2 domain of STAT3. One study screened 182,455 natural compounds from the ZINC15 database using molecular docking with various precision modes (HTVS, SP, and XP) [23]. The top candidates were further evaluated using MM-GBSA for binding free energy calculations, QikProp for pharmacokinetic properties, molecular dynamics simulations, and WaterMap analysis. This integrated approach identified ZINC67910988 as a potential STAT3 inhibitor with superior stability in molecular dynamics simulations, demonstrating the power of computational methods for drug discovery targeting dynamic domains.
Allosteric Inhibition strategies have emerged as promising approaches for targeting SH2 domains. While early drug discovery efforts focused on developing competitive inhibitors that directly target the phosphotyrosine binding pocket, recent approaches have explored allosteric sites that modulate SH2 domain function. For example, in SHP2, allosteric inhibitors stabilize the autoinhibited conformation by binding at the interface of the C-SH2 and PTP domains, preventing the conformational transitions required for activation [41]. Similar strategies may be applicable to STAT3 SH2 domain, particularly given the conformational heterogeneity observed in related SH2 domains.
Research Reagent Solutions
Table 4: Essential Research Reagents for Studying SH2 Domain Dynamics
| Reagent/Category | Specific Examples | Function/Application |
|---|---|---|
| Expression Constructs | Full-length BTK, STAT3 SH2 domain, Drk-SH2 | Protein production for structural and biophysical studies |
| Peptide Libraries | X5YX5, pTyrVar, X11 random library | Profiling SH2 domain binding specificity and affinity |
| Stabilization Additives | Non-detergent sulfobetaine (NDSB-195) | Enhance protein stability for structural studies |
| Computational Tools | ProBound, Schrӧdinger Suite, GROMACS | Modeling binding specificity and molecular dynamics |
| Display Systems | Bacterial peptide display with enzymatic phosphorylation | High-throughput affinity selection and sequencing |
Signaling Pathways and Workflow Diagrams
STAT3 Activation Pathway
Integrated Workflow for SH2 Domain Analysis
The investigation of high domain mobility and conformational heterogeneity in STAT SH2 domains represents a frontier in understanding cellular signaling and developing targeted therapeutics. The dynamic nature of these domains, once considered a challenge for structural characterization, is now recognized as fundamental to their function in phosphotyrosine-mediated signaling networks. Integrative approaches combining cryo-EM, NMR, MD simulations, and high-throughput binding assays have revealed unprecedented insights into the conformational landscapes of SH2 domains and their allosteric regulation.
Moving forward, targeting the dynamic properties of STAT SH2 domains offers promising avenues for therapeutic intervention, particularly through allosteric modulation that exploits conformational states rather than directly competing with phosphotyrosine binding. As methods for studying protein dynamics continue to advance, particularly in the areas of time-resolved structural biology and machine learning-assisted analysis of simulation data, our ability to understand and manipulate these dynamic domains will undoubtedly expand, opening new possibilities for targeting STAT signaling in disease.
Balancing Simulation Timescales with Computational Cost
The study of STAT (Signal Transducers and Activators of Transcription) proteins is pivotal for understanding cellular signaling, immune response, and cancer biology. Central to their function is the Src Homology 2 (SH2) domain, a module of approximately 100 amino acids that specifically recognizes and binds to phosphotyrosine (pY) motifs, facilitating STAT dimerization, nuclear translocation, and transcriptional activity [47] [9]. Investigating the flexibility and dynamics of the STAT SH2 domain through Molecular Dynamics (MD) simulations provides atomic-level insights into these processes. However, a significant challenge in this field is balancing the need for biologically relevant simulation timescales with the substantial computational cost this entails. This guide details advanced strategies and methodologies to navigate this trade-off, enabling more efficient and insightful research into STAT SH2 domain dynamics.
The STAT SH2 Domain: Structure and Function
The SH2 domain adopts a conserved fold comprising a central three-stranded antiparallel beta-sheet flanked by two alpha helices, forming an αβ sandwich structure [9]. A deep pocket within the βB strand contains a nearly invariant arginine residue that forms a critical salt bridge with the phosphotyrosine of peptide ligands [9]. STAT-type SH2 domains are a distinct subgroup, characterized by the absence of βE and βF strands and a split αB helix, which is an adaptation that facilitates the dimerization required for STAT transcriptional function [9].
The primary role of the SH2 domain in canonical STAT signaling is to mediate phosphotyrosine-dependent dimerization. In the cytoplasm, unphosphorylated STATs (uSTATs) await activation. Following cytokine stimulation, Janus Kinases (JAKs) phosphorylate specific tyrosine residues on receptor tails. The STAT SH2 domain then docks onto these pY sites, leading to the STAT's own phosphorylation. Subsequently, reciprocal SH2-pY interactions between two STAT monomers form an active dimer that translocates to the nucleus to regulate gene expression [47].
Table 1: Key Functional Regions of the STAT SH2 Domain
| Structural Region | Key Functional Role | Implication for Dynamics |
|---|---|---|
| pY-Binding Pocket | Binds phosphorylated tyrosine via a conserved arginine; essential for dimerization. | Simulations must capture conformational changes during ligand binding and release. |
| N-Terminal Domain | Facilitates weak dimerization of unphosphorylated STATs. | Contributes to basal-state dynamics and pre-dimerization. |
| Loop Regions (e.g., EF, BG) | Determines binding specificity for pY+3/+5 residues. | High flexibility; requires extensive sampling to understand selectivity. |
| Dimer Interface | Surface for reciprocal SH2-pY interaction in activated STATs. | Dynamics are key to understanding dimer stability and partner selection. |
Visualizing Canonical STAT Signaling and SH2 Domain Role
The following diagram illustrates the canonical STAT signaling pathway, highlighting the critical role of the SH2 domain in activation and dimerization.
Core Challenge: The Timescale-Cost Dilemma in MD
The fundamental objective of MD is to simulate atomic motions to observe biologically relevant events. For STAT SH2 domains, these include ligand binding, loop rearrangements, and dimerization interfaces. However, these events often occur on microsecond to millisecond timescales, while classical all-atom MD simulations are typically limited to nanoseconds or microseconds due to exorbitant computational costs [48]. The requirement for extensive conformational sampling of flexible regions clashes directly with the finite resources of computing time, energy, and budget.
The relationship between system size, simulation time, and computational cost is not linear but polynomial. Simulating a system twice as large for twice as long can increase the cost by a factor of four to eight. This scaling law makes the direct simulation of full-length STAT proteins or large biological complexes over long timescales prohibitively expensive for most research groups [48] [49].
Strategic Approaches and Experimental Protocols
Enhanced Sampling and Machine Learning Potentials
To overcome the timescale bottleneck, researchers employ advanced simulation strategies. Enhanced sampling methods aim to reduce the time spent simulating thermodynamically stable states, focusing computational power on crossing energy barriers. Simultaneously, Machine Learning Interatomic Potentials (MLIPs) are revolutionizing the field by providing quantum-mechanical accuracy at a fraction of the computational cost of traditional ab initio methods.
Table 2: Quantitative Comparison of Computational Methods
| Method | Typical Timescale | Relative Computational Cost | Key Applicability to STAT SH2 |
|---|---|---|---|
| Classical All-Atom MD | Nanoseconds (ns) to Microseconds (µs) | 1x (Baseline) | Good for local flexibility and loop dynamics. |
| Gaussian Accelerated MD (GaMD) | Microseconds (µs) to Milliseconds (ms) | 5-20x | Excellent for capturing large conformational changes and ligand binding/unbinding. |
| Machine Learning IPs (e.g., MACE) | Nanoseconds (ns) to Milliseconds (ms) | 10-50x (vs QM), but much faster than QM | Near-DFT accuracy for studying phosphorylation effects and metal interactions. |
| Kinetic Monte Carlo (kMC) | Seconds (s) and beyond | Highly variable | Models infrequent events like nucleation, adapted for domain assembly. |
Protocol 4.1: Implementing an AI-Driven MD Workflow with ML-IAP-Kokkos
The ML-IAP-Kokkos interface, which integrates PyTorch-based MLIPs with the LAMMPS MD package, enables fast and scalable simulations [50].
- Environment Setup: Install LAMMPS (September 2025 release or later) with Kokkos, MPI, ML-IAP, and Python support. Ensure a Python environment with PyTorch and your trained MLIP model is available.
- Model Interface Development: Implement the
MLIAPUnifiedabstract class from LAMMPS in Python. The core functioncompute_forcesmust be defined to infer pairwise forces and energies using data (atom indices, types, displacement vectors) passed from LAMMPS. - Model Serialization: Save the instantiated model object using
torch.save(mymodel, "my_model.pt"). - LAMMPS Execution: In the LAMMPS input script, load the potential using
pair_style mliap unified my_model.ptand run the simulation with Kokkos support on GPUs for optimal performance [50].
System Simplification and Multiscale Modeling
Reducing the system's complexity is a direct way to lower computational cost. A common practice is to simulate the isolated SH2 domain rather than the full-length protein, sometimes in complex with a phosphopeptide, as demonstrated in studies of the Drk-SH2 domain [51]. This drastically reduces the number of atoms. Furthermore, implicit solvent models can be used to replace explicit water molecules, eliminating thousands of solvent atoms and the associated expensive water dynamics.
For broader context, multiscale modeling couples different levels of resolution. For example, atomistic simulations of the SH2 domain can be used to parameterize a coarse-grained model, which can then simulate the entire STAT dimer over much longer timescales, providing insights into large-scale motions and interactions [48].
Protocol 4.2: NMR-Guided MD Simulations of SH2 Domain Dynamics
This protocol leverages experimental NMR data to validate and enhance MD simulations [51].
- Sample Preparation: Express and purify the recombinant STAT SH2 domain (e.g., residues 52-157 of Drk). To prevent aggregation and stabilize the domain for long-term NMR measurements, add a 1.5-fold molar excess of the target phosphopeptide (e.g., KQLpYANEGVSR from Sevenless receptor) and a stabilizing agent like NDSB-195.
- NMR Data Collection:
- Acquire 3D triple-resonance NMR spectra (HNCA, HNCOCA, etc.) for backbone and sidechain resonance assignment.
- Collect 3D 15N-separated and 13C-separated NOESY spectra to obtain NOE-derived distance restraints for structure calculation.
- Measure longitudinal (R1) and transverse (R2) 15N relaxation rates to characterize backbone dynamics on picosecond-to-nanosecond timescales.
- Structure Calculation and Validation: Calculate an ensemble of structures using programs like CYANA or AMBER with NOE and dihedral angle restraints. Validate the ensemble against the experimental NMR data.
- MD Simulation Setup and Execution:
- Use the NMR-derived structure as the starting conformation for MD.
- Solvate the system in an explicit water box, add ions to neutralize, and use a force field like AMBER or CHARMM.
- Run multiple replicas of simulations (e.g., 50-100 ns each) and check for convergence against NMR relaxation data (R1, R2). The simulated motions should be consistent with the experimental dynamics.
Integrated Workflow for Efficient Sampling
The following diagram outlines a modern, integrated workflow that combines enhanced sampling, machine learning, and experimental data to maximize sampling efficiency while managing computational expense.
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Reagents and Resources for STAT SH2 Research
| Reagent / Resource | Function and Application | Example/Specification |
|---|---|---|
| Phosphopeptide Ligands | Used in NMR and MD simulations to stabilize the SH2 domain and study binding dynamics. | KQLpYANEGVSR (from Sevenless receptor); typical purity >95% [51]. |
| Stabilizing Additives | Prevents aggregation of isolated SH2 domains in solution for stable NMR measurements. | Non-detergent sulfobetaine NDSB-195 [51]. |
| Molecular Dynamics Software | Platform for running all-atom and enhanced sampling simulations. | LAMMPS (with ML-IAP-Kokkos), AMBER, GROMACS, NAMD [50] [51]. |
| Machine Learning Potentials | Provides accurate force fields for quantum-chemical phenomena in large systems. | Models like MACE or HIPPYNN integrated via ML-IAP-Kokkos interface in LAMMPS [50]. |
| NMR Isotope Labels | Enables backbone and sidechain resonance assignment for structural and dynamics studies. | 15N-labeled ammonium chloride and 13C-labeled glucose in expression media [51]. |
Balancing simulation timescales with computational cost is a central challenge in elucidating the flexibility and function of STAT SH2 domains. No single strategy provides a perfect solution; rather, a combined approach is essential. By leveraging machine learning potentials for accuracy and speed, applying enhanced sampling to bridge timescales, simplifying systems where appropriate, and rigorously validating simulations with experimental data, researchers can effectively navigate this trade-off. The continued development of multiscale models and high-performance computing technologies promises to further push the boundaries of what is possible, deepening our understanding of STAT signaling and accelerating the development of novel therapeutic strategies.
Optimizing Force Fields and Solvation Models for Phosphotyrosine Interactions
Src Homology 2 (SH2) domains are protein interaction modules of approximately 100 amino acids that specifically recognize and bind to phosphorylated tyrosine (pY) residues, playing a fundamental role in tyrosine kinase signaling pathways [1] [9]. In the context of STAT (Signal Transducers and Activators of Transcription) proteins, SH2 domains facilitate critical protein-protein interactions that are essential for signal transduction from cytokine receptors to the nucleus [21] [9]. The molecular dynamics and flexibility of STAT SH2 domains directly influence their function in forming transcriptionally active dimers, making accurate computational modeling of these domains a crucial research objective. A significant challenge in this field lies in developing force fields and solvation models that can precisely capture the physics of phosphotyrosine interactions, which are characterized by complex electrostatic contributions and solvation effects [52] [53]. This technical guide provides a comprehensive framework for optimizing these computational parameters to advance research on STAT SH2 domains and their roles in health and disease.
Fundamental Principles of SH2 Domain-Phosphotyrosine Interactions
Structural Basis of SH2 Domain Recognition
SH2 domains maintain a highly conserved three-dimensional structure despite sequence variation, featuring a sandwich architecture composed of a central anti-parallel β-sheet flanked by two α-helices [21] [9]. The phosphotyrosine recognition mechanism involves two key binding regions: a conserved pTyr-binding pocket containing an invariant arginine residue (βB5) that forms a salt bridge with the phosphate moiety, and a specificity-determining region that interacts with residues C-terminal to the phosphotyrosine, typically at the Y+3 position [21] [9]. STAT-type SH2 domains represent a distinct structural subclass characterized by the absence of βE and βF strands and a split αB helix, adaptations that facilitate their unique dimerization functions in transcriptional regulation [9].
Electrostatic and Solvation Challenges in Phosphotyrosine Modeling
The phosphate group on phosphotyrosine presents significant modeling challenges due to its strong negative charge (-2 at physiological pH) and consequent large electrostatic solvation effects [52] [53]. Research on the p85 subunit of phosphatidylinositol 3-kinase has demonstrated that the total electrostatic solvation energy is the dominant factor determining binding affinity with ErbB3 receptor-derived phosphotyrosyl peptides [52]. Additionally, phosphotyrosine-containing peptides often interact with intrinsically disordered protein regions (IDRs), which sample diverse conformational ensembles rather than fixed structures, further complicating their computational representation [53].
Force Field Selection and Optimization Strategies
Force Field Fundamentals for Biomolecular Simulations
In molecular modeling, a force field refers to the functional forms and parameter sets used to calculate the potential energy of a system at the atomistic level [54]. The basic functional form for molecular force fields typically includes both bonded terms (covering bond stretching, angle bending, and dihedral torsions) and nonbonded terms (describing van der Waals and electrostatic interactions):
[E{\text{total}} = E{\text{bonded}} + E{\text{nonbonded}} = (E{\text{bond}} + E{\text{angle}} + E{\text{dihedral}}) + (E{\text{electrostatic}} + E{\text{van der Waals}})]
Table 1: Comparison of Force Field Treatment of Phosphotyrosine Components
| Force Field Component | Standard Treatment | Phosphotyrosine-Specific Considerations | Recommended Optimization Approaches |
|---|---|---|---|
| Bond Stretching | Harmonic potential: (E{\text{bond}} = \frac{k{ij}}{2}(l{ij}-l{0,ij})^2) [54] | Phosphoester bond lengths and vibrational frequencies | Morse potential for enhanced accuracy at bond dissociation limits [54] |
| Electrostatics | Coulomb's law: (E{\text{Coulomb}} = \frac{1}{4\pi\varepsilon0}\frac{qi qj}{r_{ij}}) [54] | -2 charge distribution over phosphate group; polarizability effects | Charge derivation using quantum mechanical protocols; polarizable force fields (AMOEBA) [55] |
| Van der Waals | Lennard-Jones potential [54] | Altered interaction parameters for phosphate oxygens | Parameterization against quantum mechanical calculations [54] |
| Dihedral Terms | Periodic functions [54] | Enhanced flexibility around phosphoester linkage | Refined parameterization using QM torsion scans [53] |
Specialized Parameters for Phosphorylated Residues
Recent advances have enabled more accurate modeling of phosphorylated residues through specialized parameter development. For the ABSINTH implicit solvent paradigm, parameters for phosphoserine (pSer) and phosphothreonine (pThr) have been developed using a thermodynamic cycle based on proton dissociation to calculate hydration free energies for each relevant charge state [53]. Similar approaches can be adapted for phosphotyrosine parameters. The free energy of solvation ((\Delta \mu_h^{A^-})) for phosphorylated residues can be calculated using:
[\Delta \muh^{A^-} = \Delta \muh^{AH} - \Delta \mud^{AH} + \Delta \mu{pKa} - \Delta \muh^{H^+}]
where (\Delta \muh^{AH}) is the hydration free energy of the protonated acid form, (\Delta \mud^{AH}) and (\Delta \mu{pKa}) represent free energy changes from proton dissociation in gas and aqueous phases, and (\Delta \mu_h^{H^+}) is the free energy of hydration of the proton [53].
Solvation Models for Phosphotyrosine Systems
Implicit Solvent Approaches
Implicit solvent models, which replace explicit solvent molecules with a continuous dielectric medium, offer computational efficiency for studying phosphotyrosine interactions, particularly when sampling large conformational ensembles of IDRs [55] [53] [56]. These models approximate the free energy of solvation ((\Delta G_{sol})) as:
[\Delta G{sol} = \Delta G{cav} + \Delta G{vdW} + \Delta G{ele}]
where the terms represent cavity formation, van der Waals interactions, and electrostatic contributions, respectively [56].
Table 2: Implicit Solvent Models for Phosphotyrosine Simulations
| Solvent Model | Theoretical Basis | Advantages for pY Systems | Limitations | Implementation Examples |
|---|---|---|---|---|
| Generalized Born (GB) | Approximation of Poisson equation using effective Born radii [56] | Computational efficiency for MD simulations; reasonable treatment of charge shielding | Less accurate for highly charged systems like pY peptides | Still et al. implementation [56]; AMBER, CHARMM |
| Poisson-Boltzmann (PB) | Numerical solution of PB equation for electrostatic potential [56] | High accuracy for electrostatic solvation of charged phosphate groups | Computationally intensive; limited compatibility with MD | APBS; DelPhi; MEAD |
| SASA Models | Solvent-Accessible Surface Area: (V{solv}^{SASA} = \sumi \sigmai^{SASA} \cdot SASAi(\vec{r_i})) [56] | Efficient modeling of nonpolar solvation contributions | Inadequate for electrostatic-dominated pY interactions | Eisenberg & McLachlan; Ooi et al. [56] |
| ABSINTH | Hybrid model combining SASA, GB, and explicit solvation for first shell [53] | Optimized for IDRs; recently parameters for pSer/pThr | Parameters for pY need validation | CAMPARI simulation engine [53] |
Explicit and Hybrid Solvent Methods
Explicit solvent models, which include individual water molecules, provide a more physically realistic representation of specific solute-solvent interactions, such as hydrogen bonding with phosphate groups and water bridging between SH2 domains and phosphopeptides [55]. Polarizable force fields like AMOEBA (Atomic Multipole Optimised Energetics for Biomolecular Applications) represent advances in explicit solvent modeling, as they account for changes in molecular charge distribution in response to environment [55]. Hybrid QM/MM (Quantum Mechanics/Molecular Mechanics) approaches offer another strategic alternative, where the phosphotyrosine and its immediate binding environment are treated with quantum mechanical methods while the remainder of the system uses molecular mechanics, providing high accuracy for the key interactions at manageable computational cost [55].
Experimental Protocols for Parameter Validation
Binding Free Energy Calculation Protocol
Surface Plasmon Resonance (SPR) analysis provides experimental binding affinities crucial for validating computational predictions [52]. The following protocol enables direct comparison between calculated and measured binding energies:
Sample Preparation: Express and purify recombinant SH2 domains. Synthesize phosphotyrosine-containing peptides corresponding to known binding motifs with purity >95%.
SPR Experimental Setup:
- Immobilize SH2 domains on CMS sensor chip using amine coupling chemistry
- Use HBS-EP buffer (10mM HEPES, 150mM NaCl, 3mM EDTA, 0.005% surfactant P20, pH 7.4) at 25°C
- Inject peptide solutions at multiple concentrations (0.1-100μM) with flow rate of 30μL/min
- Monitor association (60-120s) and dissociation (120-300s) phases
- Regenerate surface with 10mM glycine-HCl, pH 2.0
Data Analysis:
- Subtract reference cell signals and blank injections
- Fit sensograms to 1:1 Langmuir binding model to determine kinetic parameters (k(a), k(d))
- Calculate equilibrium dissociation constant K(D) = k(d)/k(_a)
- Convert to binding free energy: ΔG = -RT ln(K(_D))
Computational Validation:
- Perform molecular dynamics sampling of SH2-pY peptide complexes
- Calculate theoretical binding free energies using MM/PBSA or MM/GBSA methods
- Compare with experimental values; target correlation coefficient >0.9 as achieved in p85 N-SH2 studies [52]
Conformational Ensemble Analysis for Phosphorylated IDRs
For studying SH2 domains interacting with phosphorylated intrinsically disordered regions (IDRs), the following protocol based on all-atom Monte Carlo simulations with implicit solvent can be implemented:
System Setup:
- Obtain protein sequences from UniProt database
- Generate initial extended conformations
- Implement phosphorylated residue parameters (pSer, pThr, pTyr) in simulation package
Simulation Parameters (adapted from ABSINTH-OPLS implementation [53]):
- Use CAMPARI simulation engine with abs3.5oplsphos.prm parameter set
- Set temperature to 298K
- Perform 10(^8)-10(^9) Monte Carlo steps
- Save conformations every 10,000 steps for analysis
Analysis Metrics:
- Calculate radius of gyration (R(_g)) to assess global compaction/expansion
- Determine end-to-end distance and persistence length
- Quantify secondary structure propensities using DSSP algorithm
- Compute intramolecular contact maps
Validation Against Experimental Data:
- Compare R(_g) distributions with SAXS data
- Validate transient structural propensities with NMR chemical shifts
- Assess phosphorylation-induced conformational changes with FRET efficiency measurements
Table 3: Key Research Reagents and Computational Tools for SH2-pY Studies
| Category | Item/Resource | Specification/Function | Application Notes |
|---|---|---|---|
| Experimental Reagents | SH2 Domain Proteins | Recombinant, >95% purity, confirmed activity | STAT SH2 domains require proper folding verification |
| Phosphotyrosine Peptides | >95% purity, mass spectrometry verification | Include cognate and non-cognate sequences for specificity studies | |
| SPR Sensor Chips | CMS chips for amine coupling | Alternative: NTA chips for His-tagged proteins | |
| Computational Tools | CAMPARI | Monte Carlo simulation engine with ABSINTH implicit solvent | Optimized for IDR simulations; pSer/pThr parameters available [53] |
| AMBER/CHARMM | Molecular dynamics packages with polarizable force fields | Support for explicit solvent simulations with pY parameters | |
| APBS/DelPhi | Poisson-Boltzmann equation solvers | Accurate electrostatic calculations for binding energy decomposition | |
| Parameter Databases | MolMod Database | Force fields for molecular and ionic systems [54] | Component-specific and transferable force fields |
| openKim | Interatomic potentials database [54] | Standardized testing for force field validation |
Signaling Pathway and Methodological Framework
Diagram 1: Computational workflow for optimizing SH2-phosphotyrosine interaction models, integrating force field selection, simulation approaches, and experimental validation techniques.
Emerging Directions and Therapeutic Applications
Recent research has revealed that SH2 domains, including those in STAT proteins, can participate in the formation of biomolecular condensates through liquid-liquid phase separation (LLPS), driven by multivalent interactions [9]. This emerging understanding necessitates more sophisticated models that can capture not only specific binding interactions but also the phase behavior of SH2 domain networks. Additionally, the discovery that nearly 75% of SH2 domains interact with membrane lipids such as PIP2 and PIP3 introduces another dimension of complexity, as these interactions modulate SH2 domain function and cellular localization [9].
The therapeutic targeting of SH2 domains represents a promising approach for modulating signaling pathways in cancer and other diseases. Structure-based drug design strategies have evolved to develop inhibitors that reduce peptide character while maintaining high affinity, addressing challenges related to cell permeability and metabolic stability [21] [9]. Emerging evidence suggests that targeting lipid-binding pockets adjacent to SH2 domains or exploiting allosteric mechanisms may offer new avenues for developing selective inhibitors with improved pharmacological properties [9].
Accurate modeling of phosphotyrosine interactions with SH2 domains requires careful optimization of both force field parameters and solvation models. The strategies outlined in this guide, including specialized parameterization for phosphorylated residues, appropriate solvation model selection, and rigorous experimental validation, provide a framework for advancing research on STAT SH2 domains and their roles in cellular signaling. As computational methods continue to evolve, integration of multi-scale approaches that capture both specific molecular interactions and emergent phenomena like phase separation will be essential for fully understanding SH2 domain function and leveraging this knowledge for therapeutic development.
Strategies for Differentiating True Binding from Computational Artifacts
In the study of STAT (Signal Transducer and Activator of Transcription) proteins, their Src Homology 2 (SH2) domains are critical for phosphotyrosine-mediated signaling, dimerization, and nuclear translocation [23] [57]. For researchers investigating the molecular dynamics and flexibility of STAT SH2 domains, molecular docking and dynamics simulations are indispensable tools for identifying potential inhibitors [23]. However, a significant challenge in computational studies is the reliable distinction between true biological binding and computational artifacts—non-physiological interactions that arise from force field inaccuracies, sampling limitations, or structural biases. Such artifacts can misdirect experimental validation, wasting valuable resources. This guide provides a structured framework and practical methodologies to enhance the reliability of computational findings within STAT SH2 domain research, integrating multi-technique validation strategies suitable for researchers and drug development professionals.
Fundamental Challenges in SH2 Domain Modeling
The SH2 domain possesses a highly conserved structure—a central anti-parallel β-sheet flanked by two α-helices (the αβββα motif)—with a phosphotyrosine (pY) binding pocket divided into sub-pockets (pY+0, pY+1, pY+X) [23] [9]. This very conservation and the predominance of charged residues in the pY-binding pocket make it susceptible to certain computational artifacts.
- Electrostatic Artifacts: The pY-binding pocket contains a deeply buried, highly basic region with an invariant arginine residue (from the FLVR motif) that forms a salt bridge with the phosphate moiety of the ligand [9]. Force fields can sometimes over-stabilize these electrostatic interactions, leading to false-positive poses where a ligand is anchored primarily by its negatively charged groups without forming specific, energetically favorable van der Waals contacts or hydrogen bonds in the specificity-determining pockets.
- Structural Deformation: Long, flexible loops (e.g., the EF and BG loops) are key determinants of binding specificity by controlling access to ligand pockets [9]. During molecular dynamics (MD) simulations, these loops can undergo unrealistic deformation if the force field parameters are inaccurate, or if simulation times are too short to achieve equilibrium, resulting in an incorrect representation of the binding site's geometry and dynamics.
- Conformational Sampling: STAT-type SH2 domains, which lack the βE and βF strands found in SRC-type domains, undergo specific conformational changes upon ligand binding and dimerization [9]. Inadequate sampling of these large-scale motions in simulations can trap the system in a non-physiological conformation, preventing the observation of the true binding mode.
Integrated Methodological Framework for Validation
A single computational technique is insufficient to confirm a true binding event. The following integrated framework employs multiple methods to cross-validate results. The overall workflow for differentiating true binding from artifacts is summarized in the diagram below.
Molecular Dynamics (MD) Simulation and Stability Analysis
MD simulations are critical for assessing the stability of a docked complex under conditions mimicking the physiological environment.
- Protocol for MD Simulation:
- System Setup: Use a solvated system. A recent study on the STAT3 SH2 domain utilized the OPLS3e force field in Schrödinger's Desmond, placing the protein-ligand complex in an orthorhombic water box with TIP3P water molecules and adding ions to neutralize the system's charge [23].
- Simulation Run: Run simulations for a sufficient duration; studies on SH2 domains often use trajectories of 100 ns or longer [23]. Conduct simulations in triplicate with different initial velocities to ensure observed stability is reproducible and not due to chance.
- Stability Metrics Analysis:
- Root Mean Square Deviation (RMSD): Calculate for the protein backbone and the ligand. A complex that has reached equilibrium will show a stable RMSD plateau with fluctuations typically below 2-3 Å. Convergence of protein and ligand RMSD suggests a stable binding pose.
- Root Mean Square Fluctuation (RMSF): Analyze per-residue fluctuations. Key binding site residues should exhibit reduced fluctuations upon stable ligand binding, while flexible loops may remain dynamic.
- Intermolecular Hydrogen Bonds: Monitor the number and occupancy of specific hydrogen bonds between the protein and ligand. High-occupancy H-bonds (e.g., with key residues like Arg609, Ser611, and Ser636 in STAT3) are indicators of a specific interaction [23].
Binding Free Energy Calculations
Binding affinity calculated directly from simulation trajectories provides a more rigorous energy estimate than docking scores.
- Protocol for MM-GBSA/MM-PBSA:
- Trajectory Selection: Use a stable, equilibrated portion of the MD trajectory (e.g., the final 50-80 ns of a 100 ns simulation). Avoid using the initial, non-equilibrated phase.
- Energy Calculation: Use the Molecular Mechanics/Generalized Born Surface Area (MM-GBSA) or the Poisson-Boltzmann Surface Area (MM-PBSA) method. The Prime MM-GBSA module with the OPLS3e force field and VSGB solvation model is a common choice [23]. The binding free energy (ΔGBinding) is calculated as: ΔGBinding = GComplex - (GReceptor + GLigand).
- Interpretation: More negative values indicate stronger binding. Compare the calculated ΔGBinding for your hit compound against known inhibitors or negative controls. While absolute values may not match experiment, the relative ranking is often reliable. Per-residue energy decomposition can pinpoint which residues contribute most to binding, helping to verify the predicted binding mode.
Solvation Site Analysis
Displacing unfavorable water molecules from a binding pocket is a major driver of ligand binding. Analyzing solvation can help explain and validate binding affinities.
- Protocol for WaterMap Analysis:
- Simulation Snapshot: Use a representative structure from the MD simulation, typically an average structure or a low-energy snapshot.
- Hydration Site Identification: Employ a tool like Schrödinger's WaterMap to run molecular dynamics simulations of the solvated binding site. This identifies and characterizes hydration sites based on the density and enthalpy/entropy of water molecules [23].
- Analysis of Results: Identify "unhappy" water molecules—hydration sites with high free energy. A true binder should optimally displace these high-energy water molecules. The ligand should form enthalpically favorable interactions (e.g., H-bonds, van der Waals contacts) that surpass the free energy cost of dehydrating the binding site.
Specificity and Selectivity Profiling
A true binder should demonstrate specificity for its intended target over related domains, and its effects should be explainable within a broader biological network.
- Network Pharmacology: This computational approach maps the relationships between a compound and its multiple potential targets. Construct a compound-target network to show how a hit compound interacts not only with the STAT SH2 domain but also with related signaling pathways. This helps in visualizing polypharmacology and predicting potential off-target effects [23]. A promising, specific inhibitor should show a focused network with the primary target as a central hub.
- Experimental Specificity Models: For SH2 domains, quantitative models like ProBound can be used to predict binding affinity across the theoretical sequence space [58]. After identifying a potential ligand sequence from your compound, you can use such a model to predict its affinity for a panel of SH2 domains, providing a computational assessment of cross-reactivity risk.
Quantitative Data and Experimental Integration
The table below summarizes key metrics and their indicative values for true binding versus artifacts, derived from studies on STAT SH2 domains [23].
Table 1: Key Quantitative Metrics for Differentiating True Binding from Artifacts in STAT SH2 Domain Studies
| Metric Category | Specific Metric | Indicative of True Binding | Indicative of Computational Artifact |
|---|---|---|---|
| Simulation Stability | Protein-Ligand Complex RMSD | Plateaus and stabilizes (< 2.5 Å fluctuation) | Fails to plateau; large, continuous drift |
| Ligand-Specific Hydrogen Bonds | High occupancy (>60-70%) with key residues | Low and transient occupancy; non-specific | |
| Energetics | MM-GBSA Binding Free Energy | Significantly negative (e.g., < -40 kcal/mol) | Near zero or positive |
| Per-Residue Energy Decomposition | Major contributions from known key residues (e.g., Arg609 in STAT3) | Dominated by non-specific or surface residues | |
| Solvation | Number of Displaced Unfavorable Waters | Displacement of multiple high-energy waters | Fails to displace key unfavorable waters |
| Specificity | Network Pharmacology | Focused network with intended target as hub | Dense, promiscuous network with many off-targets |
Experimental Validation as the Ultimate Arbiter
Computational evidence must be followed by experimental validation. Several techniques are cornerstone for this:
- Empirical Affinity Measurements (SPR/ITC): Surface Plasmon Resonance (SPR) and Isothermal Titration Calorimetry (ITC) provide direct measurements of binding affinity (KD) and stoichiometry, confirming that an interaction occurs in vitro.
- Cellular Pathway Modulation: For STAT3 SH2 domain inhibitors, a key functional assay is monitoring the reduction in STAT3 phosphorylation (Y705) and its subsequent nuclear translocation, often via western blot or immunofluorescence [23] [59]. This confirms that the inhibitor disrupts the intended signaling pathway in a live cell.
- Medium/High-Throughput Phenotypic Screening: Technologies like Affimer reagents (non-antibody binding proteins) can be used to selectively target SH2 domains in intracellular assays [59]. A screen monitoring phenotypes like pERK nuclear translocation can validate the functional role of a specific SH2 domain and confirm that a small molecule inhibitor produces a similar effect.
Research Reagent Solutions Toolkit
The table below lists key reagents and computational tools essential for research in this field.
Table 2: Essential Research Reagents and Tools for STAT SH2 Domain Studies
| Reagent / Tool Name | Type | Primary Function in Research | Example Application |
|---|---|---|---|
| Affimer Proteins [59] | Protein Binding Reagent | Selective intracellular inhibition of specific SH2 domains. | Target validation; phenotypic screening (e.g., pERK nuclear translocation). |
| Schrödinger Suite (Desmond, Prime MM-GBSA, WaterMap) [23] | Computational Software Suite | Integrated platform for MD, free energy, and solvation analysis. | Assessing binding stability, affinity, and the role of water in STAT3-ligand complexes. |
| ProBound | Computational Model | Building quantitative sequence-to-affinity models for PRDs like SH2 domains. | Predicting binding affinity and specificity of peptide ligands for SH2 domains [58]. |
| ZINC15 Database | Compound Library | Public database of commercially available compounds for virtual screening. | Source of natural product libraries for in silico screening against STAT3 SH2 domain [23]. |
| Stattic & SD-36 | Small Molecule Inhibitor | Well-characterized reference inhibitors of the STAT3 SH2 domain. | Used as positive controls in functional and binding assays to benchmark new hits [23]. |
The following diagram illustrates how these computational and experimental tools integrate into a coherent strategy to conclusively identify true binders.
Differentiating true binding from computational artifacts in STAT SH2 domain research demands a rigorous, multi-faceted strategy. Relying solely on docking scores is inadequate. Instead, researchers must adopt an integrated approach that combines molecular dynamics simulations for assessing stability, advanced free energy calculations for quantifying affinity, solvation analysis for mechanistic insight, and specificity profiling for biological context. This robust computational pipeline, when systematically applied and followed by targeted experimental validation, dramatically increases the probability of identifying genuine, therapeutically relevant inhibitors of STAT SH2 domains, thereby accelerating the development of novel cancer therapeutics.
Benchmarking Insights: Validating Dynamics and Comparing Domain Families
The Src Homology 2 (SH2) domain is a structurally conserved protein module of approximately 100 amino acids that specifically recognizes and binds to phosphorylated tyrosine (pY) motifs, playing an indispensable role in intracellular signal transduction [16] [60]. Within the diverse family of SH2-containing proteins, the SH2 domains of STAT (Signal Transducer and Activator of Transcription) proteins serve a particularly critical function: they mediate the key step of STAT dimerization through reciprocal phosphotyrosine-SH2 domain interactions, which is essential for nuclear translocation and transcriptional activation [23] [61]. Unlike many other SH2 domains that primarily facilitate transient signaling complexes, STAT SH2 domains engage in stable dimerization that defines their activation cycle. Research into STAT SH2 domains is therefore not only fundamental to understanding cytokine signaling but also presents significant therapeutic opportunities, particularly in cancer and inflammatory diseases where STAT signaling is frequently dysregulated [23] [61].
Investigating the molecular dynamics and flexibility of STAT SH2 domains requires a multidisciplinary approach that integrates structural, biophysical, and computational techniques. X-ray crystallography provides high-resolution snapshots of atomic structures, Nuclear Magnetic Resonance (NMR) spectroscopy reveals dynamics and transient states in solution, and biochemical binding assays quantify interaction strengths and specificities [16] [62] [23]. Together, these methods form a complementary experimental framework that allows researchers to correlate static structures with dynamic behavior, ultimately enabling the rational design of targeted therapeutics that can modulate STAT function by disrupting pathogenic SH2 domain interactions [23].
Structural Foundations from Crystallography
X-ray crystallography has been instrumental in elucidating the canonical architecture of SH2 domains and the molecular basis of phosphopeptide recognition. The fundamental SH2 fold consists of a central anti-parallel β-sheet flanked by two α-helices, forming an αβββα structure that creates a specialized binding surface for phosphorylated tyrosine residues [16] [60] [63]. The phosphotyrosine binding pocket is located within the βB strand and features a highly conserved arginine residue at position βB5 (part of the "FLVR" motif) that forms a critical salt bridge with the phosphate moiety of the phosphorylated tyrosine [16] [63]. Additional specificity is conferred by a neighboring pocket that typically accommodates the amino acid at the +3 position C-terminal to the phosphotyrosine, creating a "two-pronged plug" binding mechanism [63].
STAT SH2 domains exhibit distinctive structural characteristics that differentiate them from prototypical Src-family SH2 domains. Specifically, STAT-type SH2 domains lack the βE and βF strands found in Src-type domains and feature a split αB helix, structural adaptations believed to facilitate the dimerization process essential for STAT activation [16]. This structural divergence highlights how evolutionary specialization of the conserved SH2 fold has yielded distinct functional capabilities.
Table 1: Key Structural Features of STAT SH2 Domains Revealed by Crystallography
| Structural Element | Description | Functional Role |
|---|---|---|
| Central β-sheet | Three-stranded anti-parallel β-sheet (βB-βD) | Scaffold for phosphotyrosine binding pocket |
| FLVR motif | Highly conserved motif with arginine at βB5 | Direct coordination of phosphotyrosine phosphate group |
| Specificity pocket | Formed by αB helix, βG strand, and loops | Recognition of residue at +3 position C-terminal to pY |
| BG and EF loops | Variable loops of differing lengths | Control access to specificity pockets; determine binding selectivity |
| Dimerization interface | Reciprocal pY-SH2 domain interaction | Mediates STAT dimerization and activation |
The application of crystallography to STAT SH2 domains has directly enabled structure-based drug design campaigns. For instance, the STAT3 SH2 domain structure revealed three distinct sub-pockets designated as pY+X (hydrophobic side), pY+0 (binds pY705), and pY+1 (binds L706), which provide complementary surfaces for targeted inhibitor development [23]. Small molecules like Stattic and SD36 were developed to bind these pockets and disrupt STAT3 dimerization, demonstrating how crystallographic data can be translated into therapeutic candidates [23].
Probing Dynamics and Flexibility through NMR Spectroscopy
While crystallography provides high-resolution structural snapshots, NMR spectroscopy offers unique insights into the dynamic behavior and conformational flexibility of SH2 domains in solution. NMR is particularly valuable for characterizing interdomain mobility and transient interactions that may be crystallographically invisible but functionally important for SH2-mediated signaling.
Studies of Src-family kinases have demonstrated the power of NMR for elucidating SH2 domain dynamics. In Fyn kinase, residual dipolar coupling and rotational diffusion anisotropy measurements revealed significant coupling yet maintained flexibility between the SH3 and SH2 domains in their peptide-bound state [62]. This interdomain flexibility has regulatory implications, as a substantial domain rearrangement is required to transition from the active state to the autoinhibited conformation where the SH2 domain engages the phosphorylated C-terminal tail [62]. Similar conformational dynamics likely govern the activation cycle of STAT proteins, where transitions between monomeric and dimeric states depend on phosphorylation status and SH2 domain accessibility.
NMR chemical shift perturbation analysis has emerged as a sensitive method for mapping binding interfaces and allosteric networks within SH2 domains. By monitoring changes in chemical shifts upon ligand binding or mutagenesis, researchers can identify residues involved in direct molecular recognition and those affected through secondary or allosteric mechanisms [62] [63]. This approach is particularly valuable for characterizing non-canonical binding modes, such as the recently discovered lipid-binding capabilities of approximately 75% of SH2 domains, including those in STAT proteins [16]. These membrane interactions, mediated by cationic regions near the phosphotyrosine-binding pocket, may modulate SH2 domain function by influencing membrane localization or altering binding kinetics [16].
For STAT SH2 domains specifically, NMR can illuminate the dynamic processes underlying dimerization and DNA binding. The transition from cytoplasmic monomers to nuclear dimers involves substantial conformational changes that can be tracked through NMR relaxation measurements and paramagnetic relaxation enhancement experiments. These techniques can quantify timescales of motion and identify transiently populated states that might represent therapeutic targets for stabilizing inactive conformations.
Quantitative Binding Analysis via Biochemical Assays
Biochemical assays provide essential quantitative parameters that complement structural and dynamic studies of STAT SH2 domains. Isothermal titration calorimetry (ITC) and surface plasmon resonance (SPR) are widely employed to determine binding affinities (Kd), stoichiometry, and thermodynamic parameters (ΔH, ΔS) for SH2 domain-phosphopeptide interactions [16]. These measurements typically reveal moderate binding affinities in the 0.1-10 μM range, which supports the physiological requirement for reversible, regulated interactions in signaling pathways [16] [9].
The development of fluorescence polarization/anisotropy assays has enabled high-throughput screening of SH2 domain inhibitors by measuring changes in molecular rotation upon binding of fluorescently labeled peptides. This approach was instrumental in identifying natural compounds that target the STAT3 SH2 domain, with candidates like ZINC67910988 demonstrating favorable binding characteristics and stability in subsequent analyses [23].
Table 2: Biochemical Assays for Characterizing STAT SH2 Domain Interactions
| Assay Method | Measured Parameters | Applications in STAT SH2 Research |
|---|---|---|
| Isothermal Titration Calorimetry (ITC) | Kd, ΔG, ΔH, ΔS, stoichiometry | Thermodynamic characterization of phosphopeptide binding |
| Surface Plasmon Resonance (SPR) | Kd, kon, koff, affinity constants | Kinetic analysis of dimerization and inhibitor interactions |
| Fluorescence Polarization | Kd, high-throughput screening | Rapid screening of compound libraries for SH2 inhibitors |
| Molecular Mechanics/Generalized Born Surface Area (MM-GBSA) | Computational binding free energy | Post-docking refinement and virtual screening prioritization |
Advanced computational methods have augmented experimental biochemical approaches. The Molecular Mechanics/Generalized Born Surface Area (MM-GBSA) method combines molecular mechanics calculations with solvation models to compute binding free energies, enabling virtual screening of potential inhibitors before experimental validation [23]. When applied to STAT3 SH2 domain inhibitors, this approach helped identify natural compounds with superior binding characteristics, such as ZINC255200449, ZINC299817570, and ZINC31167114, which demonstrated stable binding modes in subsequent molecular dynamics simulations [23].
Integrated Workflows for Comprehensive Characterization
A powerful paradigm for studying STAT SH2 domains involves the integration of multiple experimental techniques into coordinated workflows that leverage their complementary strengths. Molecular dynamics (MD) simulations serve as a particularly valuable integrator, using crystallographic structures as initial coordinates and incorporating experimental constraints from NMR and biochemical data to generate dynamic models of SH2 domain behavior [64] [23].
A representative integrated workflow might proceed as follows:
- Structural Determination: Obtain high-resolution crystal structures of STAT SH2 domains in complex with phosphopeptides or small-molecule inhibitors.
- Dynamics Characterization: Employ NMR spectroscopy to probe solution-phase dynamics and identify flexible regions.
- Binding Quantification: Use biochemical assays to measure binding affinities and kinetics for wild-type and mutant SH2 domains.
- Computational Integration: Perform molecular dynamics simulations incorporating experimental constraints to model conformational transitions and binding pathways.
- Functional Validation: Test predictions from integrated models using cellular assays of STAT signaling and transcriptional activity.
This integrative approach was exemplified in a recent study of JAK1 activation, where MD simulations revealed that bisphosphorylation of Y1034 and Y1035 in the activation loop promotes conformational transition to an open state by increasing negative charge on the tyrosine kinase domain surface and weakening its interaction with the FERM domain [64]. These simulations, informed by structural data, provided mechanistic insights that would have been difficult to obtain through any single technique.
Diagram 1: Integrated experimental workflow for STAT SH2 domain characterization. Combined approaches yield comprehensive dynamic models that enable rational drug design.
Research Reagent Solutions for STAT SH2 Domain Studies
Advancing research on STAT SH2 domains requires specialized reagents and tools that enable precise manipulation and measurement of domain structure and function. The table below outlines essential research reagents and their applications in experimental studies of STAT SH2 domains.
Table 3: Essential Research Reagents for STAT SH2 Domain Investigations
| Reagent Category | Specific Examples | Research Applications |
|---|---|---|
| Expression Constructs | His-tagged STAT SH2 domains; GST-fusion proteins | Recombinant protein production for structural and biophysical studies |
| Phosphopeptide Ligands | pY705-containing STAT3 peptides; optimized high-affinity sequences | Binding studies, specificity profiling, and competition assays |
| Small-Molecule Inhibitors | Stattic, SD36, ZINC67910988 | Functional perturbation, therapeutic development, and binding site characterization |
| NMR Isotope Labeling | 15N/13C-labeled SH2 domains; selective amino acid labeling | Backbone assignment, structure determination, and dynamics measurements |
| Crystallization Reagents | High-purity SH2 domain proteins; optimized crystallization screens | Structure determination of apo and ligand-bound states |
| Biological Samples | Phosphorylated STAT proteins; cell lysates with activated STATs | Validation of physiological relevance and cellular context studies |
These reagents enable the implementation of the integrated experimental workflows described in previous sections. For example, isotope-labeled SH2 domains permit detailed NMR investigations of dynamics, while high-purity protein preparations are essential for both crystallographic studies and quantitative biochemical assays. The development of "superbinder" SH2 domains with enhanced affinity for phosphotyrosine has further expanded the experimental toolbox, enabling applications in protein engineering and molecular trapping [60].
Emerging Techniques and Future Directions
The experimental correlates for studying STAT SH2 domains continue to evolve with technological advancements. Cryo-electron microscopy (cryo-EM) is increasingly applied to large signaling complexes involving STAT proteins, providing structural insights for complexes that may be challenging to crystallize [61]. Recent cryo-EM structures of full-length JAK kinases have revealed the spatial organization of SH2 domains within these multi-domain proteins, offering new perspectives on regulatory mechanisms [64] [61].
The role of liquid-liquid phase separation (LLPS) in SH2 domain-mediated signaling represents another emerging frontier. Multivalent interactions involving SH2 and other modular domains can drive the formation of membrane-less intracellular condensates that enhance signaling efficiency [16]. In T-cell receptor signaling, interactions among GRB2, Gads, and the LAT receptor contribute to phase-separated condensate formation that enhances signaling output [16]. Similar mechanisms may operate in STAT signaling pathways, suggesting new dimensions of SH2 domain organization beyond binary interactions.
Advanced computational methods are also expanding the correlates of experimental observation. Extended molecular dynamics simulations (reaching microsecond timescales) provide increasingly accurate models of conformational transitions, while WaterMap analysis offers insights into the role of solvation in SH2 domain binding and inhibitor design [23]. These computational approaches, when tightly coupled with experimental validation, promise to accelerate the discovery of next-generation STAT SH2 domain inhibitors with improved potency and selectivity.
Diagram 2: Emerging research directions and therapeutic applications for STAT SH2 domain studies. New methodological approaches are expanding understanding and enabling novel therapeutic interventions.
The comprehensive understanding of STAT SH2 domain function requires the integration of multiple experimental correlates that span resolution from atomic structure to cellular context. Crystallography provides the essential structural framework, NMR reveals dynamic properties in solution, biochemical assays quantify interaction parameters, and computational methods integrate these data into predictive models. This multidisciplinary approach has illuminated not only the canonical phosphotyrosine recognition function of STAT SH2 domains but also emerging roles in membrane interactions, phase separation, and allosteric regulation.
As technical capabilities advance, particularly in cryo-EM, molecular simulations, and high-throughput screening, the experimental correlates for STAT SH2 domain research will continue to evolve and refine our understanding of these critical signaling modules. These advances will undoubtedly accelerate the development of targeted therapeutics for cancer, inflammatory diseases, and immune disorders where STAT signaling plays a central pathogenic role. The integrated application of structural, biophysical, and computational methods thus represents the most promising path forward for both basic science and translational applications targeting STAT SH2 domains.
Src Homology 2 (SH2) domains are crucial protein modules that direct cellular signaling by specifically recognizing phosphotyrosine (pY) motifs, thereby orchestrating processes such as cell growth, differentiation, and immune responses [9] [21]. Despite a conserved core structure, SH2 domains exhibit significant functional and structural divergence. Two major evolutionary groups are the STAT-type and Src-type SH2 domains [10] [6]. STAT-type SH2 domains are found in Signal Transducers and Activators of Transcription proteins and are characterized by a C-terminal α-helix (αB'). In contrast, Src-type SH2 domains, present in Src family kinases (SFKs) like Hck and c-Src, typically feature a C-terminal β-sheet (βE-βF) [10] [6]. This structural variation profoundly influences their conformational flexibility, mechanisms of regulation, and ultimately, their potential as drug targets. Framed within a broader thesis on molecular dynamics, this analysis examines how the inherent flexibility of STAT SH2 domains, compared to their Src-family counterparts, creates unique challenges and opportunities for therapeutic intervention.
Structural Classification and Functional Implications
The canonical SH2 domain fold consists of a central anti-parallel β-sheet flanked by two α-helices, forming an αββββα motif [9] [16]. The key to phosphopeptide binding lies in two sub-pockets: a pTyr pocket that engages the phosphorylated tyrosine, and a pY+3 pocket that confers specificity by recognizing residues C-terminal to the pY [10] [9]. The critical structural divergence between STAT and Src-type SH2 domains occurs in the C-terminal region following the αB helix.
Table 1: Fundamental Classification of SH2 Domain Types
| Feature | STAT-Type SH2 Domains | Src-Type SH2 Domains |
|---|---|---|
| Defining C-Terminal Structure | C-terminal α-helix (αB') [10] [6] | C-terminal β-sheet (βE and βF strands) [10] [6] |
| Representative Proteins | STAT1, STAT3, STAT5 [10] [16] | Src, Hck, Fyn, Lck [65] [9] [16] |
| Primary Functional Role | Mediate STAT dimerization and nuclear translocation for gene transcription [10] | Mediate autoinhibition and recruitment to signaling complexes; regulate kinase activity [65] [21] |
This structural difference is not merely topological. The αB' helix in STAT SH2 domains participates in critical cross-domain interactions that are essential for STAT dimerization and transcriptional function [10]. In Src-family kinases, the SH2 domain plays a key autoinhibitory role by engaging the phosphorylated C-terminal tail and the SH2-kinase linker, thereby stabilizing the kinase in a closed, inactive conformation [65] [21].
Figure 1: Structural and Functional Classification of SH2 Domains
Comparative Analysis of Flexibility and Dynamics
Conformational Flexibility in STAT SH2 Domains
STAT SH2 domains exhibit significant inherent flexibility, which is crucial for their function. Molecular dynamics simulations and structural studies reveal that the pY pocket of STAT SH2 domains is highly dynamic, with its accessible volume varying dramatically even on sub-microsecond timescales [10]. This flexibility is a critical consideration for drug discovery, as crystal structures may not capture the domain in a state conducive to inhibitor binding [10]. Furthermore, the BC* loop and the αB' helix are involved in both phosphopeptide binding and STAT dimerization, implying that residues in the pY+3 pocket can exert dual effects on these processes [10]. This interconnectedness suggests that ligand binding can allosterically influence dimerization, and vice versa.
Regulatory Flexibility in Src Family SH2 Domains
The flexibility of Src-family SH2 domains is context-dependent and integral to kinase regulation. In the down-regulated state, the SH2 domain engages in a rigid, intramolecular interaction with the phosphorylated C-terminal tail. However, studies on Hck reveal that its SH2-kinase linker is a suboptimal ligand for the isolated SH3 domain, adopting a stable polyproline type II (PPII) helix only within the context of the full-length, autoinhibited protein [65]. This creates a "conformational switch" where the SH2 and SH3 domains work in concert, making the kinase uniquely sensitive to activation by external SH3-binding proteins like HIV-1 Nef [65]. This highlights a form of allosteric flexibility where the stability of the inactive conformation is fine-tuned and can be disrupted by specific intermolecular interactions.
Table 2: Flexibility and Dynamics of SH2 Domains
| Characteristic | STAT-Type SH2 Domains | Src-Type SH2 Domains |
|---|---|---|
| Inherent Conformational Dynamics | High; pY pocket is highly flexible with large volume variations [10] | More constrained in the autoinhibited state; stability is context-dependent [65] |
| Key Flexible Elements | pY pocket, BC* loop, αB' helix [10] | SH2-kinase linker, which is a suboptimal SH3 ligand [65] |
| Functional Implication of Flexibility | Allosteric linkage between peptide binding and dimerization; challenge for static drug design [10] | Forms a "conformational switch" for kinase regulation; sensitive to activation by SH3 ligands [65] |
Druggability and Therapeutic Targeting
The STAT SH2 Domain as a Challenging Target
STAT3 has been historically classified as "undruggable," primarily due to the challenges posed by its shallow, hydrophilic pY-binding pocket, which is designed to recognize a phosphorylated tyrosine residue [66]. High-value targets like STAT3 contribute to multiple hallmarks of cancer, creating a significant unmet medical need [66]. The inherent flexibility of the STAT SH2 domain adds a layer of complexity, as effective inhibitors must target a dynamic binding interface rather than a static, well-defined pocket.
Breakthroughs in Targeting the STAT3 SH2 Domain
A key breakthrough in targeting STAT3 came from a sophisticated virtual ligand screening (VLS) strategy. This approach was based on the observation that the STAT3 SH2 domain binds high-affinity pY-peptides in a β-turn conformation, similar to the GRB2 SH2 domain [66]. In this folded conformation, the critical residues on STAT3 that interact with the peptide are within 10 Å of each other, a distance that can be bridged by a small, drug-like molecule [66]. This insight led to the identification and optimization of TTI-101 (C188-9), a potent, oral small-molecule inhibitor that binds directly to the STAT3 SH2 domain with a Ki of 12.4 nM [66]. TTI-101 inhibits STAT3 phosphorylation, dimerization, and the proliferation of cancer cells driven by STAT3, and it is now in clinical trials for advanced solid tumors [66]. This success demonstrates that the flexibility and shallow pocket of STAT SH2 domains can be overcome with precise structural insights.
Figure 2: Workflow for Developing the STAT3 SH2 Inhibitor TTI-101
Targeting Src Family SH2 Domains
In contrast, targeting Src-family SH2 domains has faced different hurdles. The primary challenge has been the liability and poor cell permeability of negatively charged, phosphorylated SH2 ligand mimics [21]. Extensive structure-based drug design efforts have focused on reducing the size, charge, and peptide character of these ligands. This has led to the development of high-affinity lead compounds for Grb2 and Src SH2 domains with potent cellular activity [21]. The more well-defined, two-pronged "plug-and-socket" binding mode of Src-type SH2 domains offers a somewhat more classical structure-based drug design path, albeit one still complicated by the physicochemical properties of phosphate mimics.
Table 3: Druggability and Therapeutic Targeting Landscape
| Aspect | STAT-Type SH2 Domains | Src-Type SH2 Domains |
|---|---|---|
| Historical Classification | "Undruggable" [66] | Challenging, but druggable [21] |
| Primary Targeting Challenge | Shallow, flexible, hydrophilic pY pocket [66] | Peptidic, charged ligands with poor drug-like properties [21] |
| Key Targeting Strategy | Virtual screening based on β-turn peptide conformation; allosteric inhibition [66] | Structure-based design to create non-peptidic, cell-permeable phosphate mimics [21] |
| Clinical Stage Inhibitor | TTI-101 (STAT3 inhibitor, Phase 1) [66] | Various leads (e.g., for Grb2, Src); extensive pre-clinical development [21] |
Experimental Approaches and Research Toolkit
A multi-faceted approach is required to study the flexibility and druggability of SH2 domains. Key experimental methodologies and reagents are detailed below.
Table 4: The Scientist's Toolkit for SH2 Domain Research
| Method/Reagent | Function/Description | Key Application |
|---|---|---|
| X-ray Crystallography | Determines high-resolution 3D atomic structures of proteins and complexes. | Solved structures of down-regulated Hck and STAT SH2 domains, revealing autoinhibitory mechanisms and structural variations [65] [10]. |
| Molecular Dynamics (MD) Simulations | Computational method simulating physical movements of atoms over time. | Used to analyze JAK1 TK domain dynamics and the effect of phosphorylation on conformational change, relevant to SH2-associated kinases [64]. |
| Oriented Peptide Array Library (OPAL) | High-throughput screening to define the binding specificity and motif of SH2 domains. | Used to define the specificity space of 76 human SH2 domains, enabling prediction of binding partners [67]. |
| Surface Plasmon Resonance (SPR) | Label-free technique to measure biomolecular binding kinetics and affinity. | Used to determine the binding affinity (Ki=12.4 nM) of TTI-101 for the STAT3 SH2 domain [66]. |
| Virtual Ligand Screening (VLS) | Computational docking of compound libraries into protein structures to identify hits. | Identified initial STAT3 SH2 inhibitors from 920,000 compounds, leading to TTI-101 [66]. |
| In-vitro Kinase Assay (e.g., Z'-Lyte) | Biochemical assay to measure kinase activity and inhibition. | Used to measure Hck and Src activation induced by SH3-binding peptides [65]. |
Figure 3: Experimental Workflow for SH2 Domain Research
The comparative analysis of STAT and Src-family SH2 domains reveals a fundamental trade-off between structural flexibility and druggability. STAT-type SH2 domains are characterized by high conformational dynamics, which is critical for their allosteric regulation and function in transcription. This very flexibility, however, has historically made them appear "undruggable." The success of TTI-101 proves this barrier can be overcome by leveraging deep structural insights, such as the β-turn binding mode, to design effective small-molecule inhibitors. In contrast, Src-type SH2 domains exhibit a more context-dependent flexibility that is central to their role as conformational switches in kinase regulation. Their primary druggability challenge lies in the physicochemical properties of their ligands rather than an intrinsically dynamic architecture. Future research, leveraging advanced techniques like long-timescale molecular dynamics and integrative structural biology, will continue to decode the allosteric networks within these domains. This will accelerate the rational design of next-generation inhibitors that can precisely modulate the flexibility of SH2 domains to treat cancer, immune disorders, and other diseases.
The Src Homology 2 (SH2) domain is a critical protein-protein interaction module found in numerous signaling proteins, specializing in recognizing and binding sequences containing phosphorylated tyrosine residues [1] [9]. In the context of STAT (Signal Transducer and Activator of Transcription) proteins, the SH2 domain mediates key interactions essential for their activation and function, particularly through facilitating dimerization via phosphotyrosine-SH2 domain binding [23]. Given the central role of STAT proteins, especially STAT3, in cancer progression and immune evasion, the SH2 domain has emerged as a promising therapeutic target [9] [23].
Evaluating inhibitor efficacy against this domain requires robust biophysical and computational methods. This technical guide focuses on two powerful approaches: binding free energy calculations, which provide atomic-level insights into inhibitor interactions through computational simulations, and thermal shift assays, which experimentally measure compound-induced changes in protein stability. When applied to STAT SH2 domains, these methods offer complementary data critical for advancing molecular dynamics research and structure-based drug discovery.
Binding Free Energy Calculations
Binding free energy calculations are computational techniques that predict the strength of interaction between a protein and a ligand by estimating the free energy change (ΔG) upon binding. Accurate prediction of binding affinities is crucial for rational drug design as it directly correlates with inhibitor potency [68].
Theoretical Background and Methodologies
Free Energy Perturbation (FEP)
Free Energy Perturbation is an alchemical method that calculates the free energy difference between two states by gradually transforming one ligand into another within the binding site. The relative binding free energy (ΔΔG) between ligand A and B is calculated using the thermodynamic cycle shown in Figure 1, where ΔΔGA→B = ΔGBbind - ΔGAbind = ΔGA→Bp - ΔGA→Bw [68].
The FEP calculations utilize a mapping potential defined as: εm(λm) = U1(1 - λm) + U2λm where U1 and U2 represent the potential energies of the initial and final states, and λm is the mapping parameter that varies from 0 to 1 [68].
For transformations involving creation or annihilation of atoms, a modified soft-core Lennard-Jones potential is employed to avoid sampling issues: UijLJ(rij;λ) = λ(Bi×Bj)2(Ai×Aj)(1/[α(1-λ)2 + rij6((Bi×Bj)/(Ai×Aj))]2 - 1/[α(1-λ)2 + rij6((Bi×Bj)/(Ai×Aj))]) [68]
Molecular Mechanics Generalized Born Surface Area (MM-GBSA)
MM-GBSA is an end-point method that calculates binding free energy using the equation: ΔGBinding = ΔGComplex - (ΔGReceptor + ΔGLigand) where more negative values indicate stronger binding [23]. This method combines molecular mechanics calculations with implicit solvation models and is computationally efficient for screening large compound libraries [23].
Table 1: Comparison of Binding Free Energy Calculation Methods
| Method | Theoretical Basis | Computational Cost | Accuracy | Best Use Cases |
|---|---|---|---|---|
| FEP | Alchemical transformations with thermodynamic cycle | High (requires extensive sampling) | High (within 1 kcal/mol) | Lead optimization, relative affinity predictions [68] |
| MM-GBSA | End-point sampling with implicit solvation | Moderate | Moderate | Virtual screening, binding mode analysis [23] |
| Replica Exchange FEP | Enhanced sampling with replica exchange | Very High | Similar to FEP (limited by force field) | Systems with slow conformational changes [68] |
Application to STAT SH2 Domain Research
For STAT SH2 domains, these methods have been successfully applied to identify and optimize inhibitors. A recent study screened 182,455 natural compounds against the STAT3 SH2 domain using molecular docking followed by MM-GBSA calculations to identify promising inhibitors that disrupt STAT3 dimerization [23]. Key interaction residues in the STAT3 SH2 domain include Arg609, Glu594, Lys591, Ser636, Ser611, Val637, Tyr657, Gln644, Thr640, Glu638, and Trp623, which form hydrogen bonds and hydrophobic interactions with inhibitors [23].
The SH2 domain structure consists of an αβββα motif with a central anti-parallel β-sheet flanked by two α-helices [23]. The phosphotyrosine (pY) binding pocket is divided into three sub-pockets: pY+X (hydrophobic side), pY+0 (binds pY705), and pY+1 (binds L706) [23]. Understanding these structural features is essential for interpreting binding free energy calculations.
Figure 1: Workflow for computational screening of STAT3 SH2 domain inhibitors, from compound preparation through binding affinity assessment [23].
Thermal Shift Assays
Thermal Shift Assay (TSA), also known as differential scanning fluorimetry (DSF), measures changes in protein thermal stability induced by ligand binding [69]. When a ligand binds to its target protein, it often stabilizes the protein's native structure, resulting in an increased thermal denaturation temperature (Tm or Tagg) [69] [70].
Experimental Principles and Methodologies
The underlying principle of TSA is that small molecule binding can increase the thermal stability of a protein by shifting the equilibrium between native and denatured states toward the native form [69]. This stabilization effect is quantified by measuring the temperature at which the protein unfolds, providing information about target engagement and binding affinity [70].
nanoDSF (nano Differential Scanning Fluorimetry)
nanoDSF relies on the intrinsic fluorescence of tryptophan or tyrosine residues in proteins. As the protein unfolds, these residues become exposed to solvent, resulting in a shift of fluorescence emission wavelength from approximately 330 nm to 350 nm [69]. The protein must contain intrinsically fluorescent residues, typically tryptophan or tyrosine, for this label-free method [69].
Thermofluor
This method utilizes extrinsic fluorogenic dyes such as SYPRO Orange, which binds nonspecifically to hydrophobic surfaces [69]. In its unbound state, the dye's fluorescence is quenched by water, but when the protein unfolds and exposes hydrophobic regions, the dye binds and fluoresces [69]. This method is compatible with standard qPCR machines and allows high-throughput screening [69].
Cellular Thermal Shift Assay (CETSA)
CETSA extends thermal shift principles to cellular environments, providing evidence of target engagement in a more physiologically relevant context [70]. High-throughput CETSA (HT-CETSA) formats have been developed using various detection methods, including AlphaLISA assays and nanoluciferase reporters [70].
Table 2: Thermal Shift Assay Methods and Their Characteristics
| Method | Detection Mechanism | Throughput | Sample Type | Key Advantages |
|---|---|---|---|---|
| nanoDSF | Intrinsic tryptophan/tyrosine fluorescence | Medium | Purified protein | Label-free, no dye required [69] |
| Thermofluor | Extrinsic dye (SYPRO Orange) binding | High | Purified protein | Compatible with qPCR instruments [69] |
| CPM Assay | Thiol-specific dye fluorescence | Medium | Purified protein | Effective for membrane proteins [69] |
| CETSA | Antibody-based or MS detection | Medium to High | Cells or lysates | Cellular context, endogenous protein [70] |
| HT-CETSA | AlphaLISA, nanoluciferase reporters | Very High | Cells or lysates | Suitable for compound screening [70] |
Experimental Protocol for STAT SH2 Domain TSA
A typical thermal shift assay for evaluating SH2 domain inhibitors includes the following steps [69]:
Materials Preparation:
- Fluorometer with temperature control or qPCR machine
- Suitable fluorescent dye (SYPRO Orange for Thermofluor; intrinsic fluorescence for nanoDSF)
- Assay plate (96-well or 384-well qPCR plate)
- Test compounds at 50- to 100-fold concentrated solutions (typically 10-100 mM)
- Target protein (STAT SH2 domain) diluted to working concentration (0.5-5 μM) in assay buffer
Assay Procedure:
- Dispense protein solution with dye into assay plates
- Add test compounds to appropriate wells (include DMSO-only negative controls)
- Centrifuge plates briefly (~1000 × g, 1 min) to mix solutions
- Overlay with silicone oil or apply plastic seals to prevent evaporation
- Perform additional centrifugation (~1000 × g, 1 min)
- Run temperature ramp (typically 1°C/min) from 25°C to 95°C
- Measure fluorescence at regular intervals (0.2-1.0°C per reading)
Data Analysis:
- Generate melt curves by plotting fluorescence versus temperature
- Calculate melting temperature (Tm) for each condition
- Determine ΔTm (Tm protein+ligand - Tm protein alone)
- Compounds producing significant ΔTm values (typically >1°C) indicate binding
Figure 2: Thermal shift assay workflow for evaluating STAT SH2 domain inhibitors, from sample preparation through data interpretation [69].
Research Reagent Solutions
Table 3: Essential Research Reagents for SH2 Domain Studies
| Reagent/Category | Specific Examples | Function/Application | Experimental Context |
|---|---|---|---|
| Fluorescent Dyes | SYPRO Orange, CPM dye, DCVJ | Detect protein unfolding in TSA | Thermofluor assays with purified SH2 domains [69] |
| Computational Software | Schrödinger Suite, Molaris-XG | Molecular docking, FEP, MM-GBSA | Virtual screening of SH2 domain inhibitors [68] [23] |
| Protein Production | Recombinant STAT SH2 domains | Provide target for biophysical assays | Purified protein for TSA and structural studies [69] |
| Detection Systems | AlphaLISA, nanoluciferase | Detect protein in cellular assays | HT-CETSA for cellular target engagement [70] |
| Compound Libraries | Natural product databases, focused libraries | Source of potential inhibitors | Screening for novel SH2 domain binders [23] |
Integrated Application in STAT SH2 Domain Research
The combination of binding free energy calculations and thermal shift assays provides a powerful framework for evaluating STAT SH2 domain inhibitors. Computational methods offer atomic-level insights into binding mechanisms and enable rapid screening of large compound libraries, while experimental assays validate target engagement and provide quantitative measures of compound effects in relevant biological contexts [23] [70].
For STAT3 SH2 domain research, this integrated approach has identified natural compounds such as ZINC255200449, ZINC299817570, ZINC31167114, and ZINC67910988 as potential inhibitors based on their favorable binding affinities in MM-GBSA calculations and stability in molecular dynamics simulations [23]. Subsequent experimental validation using thermal shift assays can confirm the stabilizing effects of these compounds on the STAT3 SH2 domain.
The molecular dynamics and flexibility of STAT SH2 domains play a crucial role in their function and inhibitor binding. SH2 domains typically display a conserved fold consisting of a three-stranded antiparallel beta-sheet flanked by two alpha helices (αA-βB-βC-βD-αB) [9]. STAT-type SH2 domains are distinct in that they lack the βE and βF strands found in SRC-type SH2 domains, with the αB helix split into two helices - an adaptation that facilitates STAT dimerization [9]. Understanding these structural dynamics is essential for interpreting both computational and experimental data on inhibitor binding.
Recent advances in high-throughput cellular thermal shift assays (HT-CETSA) now enable target engagement studies in more physiologically relevant environments, bridging the gap between computational predictions and cellular efficacy [70]. Similarly, improvements in free energy calculation methods, including replica exchange techniques and advanced sampling algorithms, continue to enhance the accuracy of binding affinity predictions for SH2 domain inhibitors [68].
Allosteric inhibition has emerged as a powerful strategy in drug discovery, offering distinct advantages over traditional orthosteric targeting. Unlike orthosteric inhibitors that compete with substrates for the active site, allosteric inhibitors bind to topographically distinct sites, inducing conformational changes that modulate protein activity [71] [72]. This mechanism provides enhanced selectivity for closely related protein families due to lower evolutionary conservation of allosteric sites and the potential for fine-tuned modulation rather than complete inhibition [73] [71]. The therapeutic promise of this approach is exemplified by drugs like SHP099, an allosteric inhibitor of the phosphatase SHP2, which stabilizes the inactive conformation through binding at the interface of multiple domains [74].
Complementing allosteric modulation, multivalent targeting employs compounds with multiple binding moieties to engage several sites on one or more target proteins simultaneously [75]. This strategy capitalizes on avidity effects, where the combined binding strength exceeds the sum of individual interactions, resulting in dramatically increased affinity and selectivity [75] [76]. Multivalent ligands can be categorized as bivalent (engaging two orthosteric sites) or bitopic (engaging both orthosteric and secondary sites), with applications in targeting receptor oligomers and enhancing cellular internalization [75] [76].
When applied to challenging targets like the STAT3 SH2 domain, these strategies offer promising avenues for disrupting protein-protein interactions that have traditionally been difficult to drug with small molecules. The convergence of allosteric and multivalent approaches represents a frontier in therapeutic development, particularly for oncology targets where STAT3 plays a pivotal role [1] [5].
The STAT3 SH2 Domain as a Therapeutic Target
Structural and Functional Significance
The Src Homology 2 (SH2) domain is a protein module of approximately 100 amino acids that specifically recognizes and binds to phosphorylated tyrosine residues, serving as a crucial "reader" in cellular signaling networks [1]. SH2 domains are found in 111 human proteins, including kinases, phosphatases, and adaptor proteins, where they facilitate the assembly of signaling complexes in response to tyrosine phosphorylation [1].
STAT3 (Signal Transducer and Activator of Transcription 3) contains a critical SH2 domain that mediates its dimerization and activation [5]. Following phosphorylation at tyrosine 705 (Y705) by upstream kinases, STAT3 molecules engage in reciprocal SH2-pY705 interactions, forming active dimers that translocate to the nucleus and drive the expression of genes involved in cell proliferation, survival, and immune evasion [5]. This activation mechanism makes the STAT3 SH2 domain an attractive target for cancer therapy, particularly since constitutive STAT3 activation is a hallmark of numerous malignancies, including breast, prostate, lung, and hematological cancers [5].
Structural Architecture of the STAT3 SH2 Domain
The SH2 domain adopts a conserved αββα fold consisting of a central anti-parallel β-sheet flanked by two α-helices [1] [5]. The phosphotyrosine (pY) binding pocket of STAT3's SH2 domain is structurally organized into three sub-pockets:
- pY+0 pocket: Binds the phosphorylated Y705 residue
- pY+1 pocket: Accommodates the leucine at position 706 (L706)
- pY+X pocket: A hydrophobic region that engages additional residues [5]
Key residues involved in binding include Arg609, Glu594, Lys591, Ser636, Ser611, Val637, Tyr657, Gln644, Thr640, Glu638, and Trp623, which form an interaction network stabilizing the phosphoryrosine-containing motif [5]. Disrupting these interactions through allosteric or multivalent inhibition prevents STAT3 dimerization, nuclear translocation, and subsequent transcriptional activity.
Computational Approaches for Allosteric Inhibitor Discovery
Molecular Docking and Virtual Screening
Computational screening has emerged as a powerful strategy for identifying potential allosteric inhibitors of the STAT3 SH2 domain. A recent comprehensive study employed an in silico workflow to screen 182,455 natural compounds from the ZINC15 database [5]. The methodology proceeded through several stages:
Protein Preparation: The STAT3 crystal structure (PDB: 6NJS) was selected based on superior resolution (2.70 Å) and integrity of the SH2 domain. The protein structure was processed using the Protein Preparation Wizard in Schrödinger Suite, which involved adding hydrogen atoms, filling missing side chains, and energy minimization using the OPLS3e force field [5].
Ligand Preparation: Natural compounds were processed with LigPrep to generate three-dimensional structures with optimized ionization states at physiological pH (7.4 ± 0.5) [5].
Docking Protocol: The screening employed a multi-tiered approach:
- High-Throughput Virtual Screening (HTVS) of the entire library
- Standard Precision (SP) docking of the top 55,872 compounds from HTVS
- Extra Precision (XP) docking of the most promising candidates (cut-off at -6.5 kcal/mol) [5]
Binding Affinity Assessment: Molecular Mechanics-Generalized Born Surface Area (MM-GBSA) calculations were performed to determine binding free energies (ΔG Binding) using the equation:
ΔG Binding = ΔG Complex - (ΔG Receptor + ΔG Ligand) [5]
Table 1: Top Natural Compound Inhibitors of STAT3 SH2 Domain Identified Through Virtual Screening
| Compound ID | Docking Score (kcal/mol) | MM-GBSA ΔG (kcal/mol) | Key Interactions |
|---|---|---|---|
| ZINC255200449 | -10.2 | -45.8 | Arg609, Ser611 |
| ZINC299817570 | -9.8 | -43.5 | Glu594, Ser636 |
| ZINC31167114 | -9.5 | -42.1 | Lys591, Tyr657 |
| ZINC67910988 | -11.3 | -49.2 | Multiple residues |
Molecular Dynamics and Validation
Compounds exhibiting favorable binding affinities and pharmacokinetic properties underwent further validation through molecular dynamics (MD) simulations using Desmond software [5]. These simulations assessed:
- Complex stability through root-mean-square deviation (RMSD) and fluctuation (RMSF) analyses
- Interaction persistence throughout the simulation trajectory
- Solvation effects using WaterMap analysis
- Electronic properties through Density Functional Theory (DFT) calculations
The lead compound ZINC67910988 demonstrated superior stability in MD simulations, maintained key interactions with the SH2 domain, and exhibited favorable electronic properties with a well-defined HOMO-LUMO gap [5].
Multivalent Targeting Strategies for Enhanced Efficacy
Design Principles and Classification
Multivalent compounds offer significant advantages for targeting challenging protein-protein interactions like STAT3 dimerization. These constructs can be systematically categorized based on their architecture and binding mode [75]:
Table 2: Classification of Multivalent Targeting Strategies
| Category | Binding Sites Engaged | Application to STAT3 | Key Advantages |
|---|---|---|---|
| Homobivalent | Two identical orthosteric sites | Simultaneously targeting two STAT3 SH2 domains | Avidity effects, increased potency |
| Heterobivalent | Two different orthosteric sites | Targeting STAT3 SH2 domain and cooperating receptor | Enhanced selectivity for specific cellular contexts |
| Cis-bitopic | Orthosteric + allosteric sites on same protein | Engaging both pY705 binding pocket and allosteric site on STAT3 | Synergistic inhibition, novel mechanisms |
| Trans-bitopic | Sites on neighboring proteins | Bridging STAT3 with regulatory proteins | Redirecting protein interactions |
| Intercellular | Receptors on different cells | Engaging STAT3 on cancer and immune cells | Immunotherapeutic applications |
Experimental Evidence and Applications
The therapeutic potential of multivalent constructs is exemplified by their success in other signaling systems. In opioid receptor targeting, optimized homobivalent compounds demonstrated potencies that exceeded their monovalent counterparts by several orders of magnitude, with both the pharmacophore identity and linker length critically influencing activity [75]. Similarly, bitopic ligands for GPCRs have enabled biased signaling, selectively activating beneficial pathways while avoiding detrimental ones [75].
For STAT3 inhibition, multivalent approaches could simultaneously engage both the pY705 binding pocket and neighboring allosteric sites, potentially overcoming limitations of monovalent inhibitors. The development of protein-drug conjugates (PDCs) with multivalent architectures has shown enhanced tumor targeting and internalization in oncology applications, providing a blueprint for similar approaches against STAT3 [76].
Experimental Protocols for Allosteric Inhibitor Validation
Biophysical Characterization
Nuclear Magnetic Resonance (NMR) Spectroscopy
- Purpose: Map conformational dynamics and allosteric perturbations at atomic resolution
- Protocol: Acquire [¹H-¹⁵N]-TROSY-HSQC spectra of isotopically labeled STAT3 SH2 domain
- Analysis: Monitor chemical shift perturbations upon inhibitor binding
- Application: Identify residues involved in allosteric communication networks [73] [74]
Surface Plasmon Resonance (SPR)
- Purpose: Determine binding kinetics (kon, koff) and affinity (KD)
- Immobilization: Covalently immobilize STAT3 SH2 domain on CMS sensor chip
- Ligand Injection: Flow inhibitors at varying concentrations (0.1-100 × KD)
- Data Fitting: Use 1:1 binding model to extract kinetic parameters [77]
Isothermal Titration Calorimetry (ITC)
- Purpose: Measure thermodynamic parameters of binding (ΔH, ΔS, ΔG, Kd)
- Protocol: Inject inhibitor solutions into STAT3 SH2 domain in sample cell
- Conditions: 25°C, PBS buffer, matching compound and protein buffers
- Analysis: Fit integrated heat data to single-site binding model [74]
Functional and Cellular Assays
Phosphorylation Status Analysis
- Objective: Monitor Y705 phosphorylation as indicator of STAT3 activation
- Method: Western blotting with phospho-specific STAT3 (Y705) antibody
- Cell Treatment: Expose STAT3-dependent cancer cell lines to inhibitors (1-100 μM, 2-24h)
- Normalization: Total STAT3 and loading controls (β-actin/GAPDH) [5]
Dimerization Assay
- Principle: Assess inhibitor effect on STAT3 dimer formation
- Protocol: Co-immunoprecipitation or crosslinking followed by SDS-PAGE
- Quantification: Compare dimer:monomer ratios in treated vs. untreated cells [5]
Gene Expression Profiling
- Target Genes: Quantify mRNA levels of STAT3-regulated genes (Bcl-2, Cyclin D1, c-Myc)
- Methods: Quantitative RT-PCR or RNA sequencing
- Application: Determine functional consequences of SH2 domain inhibition [5]
Research Reagent Solutions
Table 3: Essential Research Tools for STAT3 SH2 Domain Studies
| Reagent/Category | Specific Examples | Research Application |
|---|---|---|
| STAT3 SH2 Domain Constructs | Recombinant human STAT3 SH2 domain (residues 575-688) | Biophysical studies, inhibitor screening |
| Reference Inhibitors | Stattic, SD-36 | Positive controls for inhibition assays |
| Antibodies | Anti-pY705-STAT3, total STAT3, secondary antibodies with fluorescent/HRP conjugates | Cellular validation, Western blotting |
| Cell Lines | MDA-MB-231 (breast cancer), DU145 (prostate cancer) | Cellular activity assessment |
| Computational Tools | Schrödinger Suite (Maestro), Desmond, PyMOL | Virtual screening, MD simulations |
| NMR Isotopes | ¹⁵N-ammonium chloride, ¹³C-glucose | Isotopic labeling for NMR studies |
Signaling Pathways and Methodological Workflows
STAT3 Activation and Allosteric Inhibition Pathway
Computational Screening Workflow for STAT3 SH2 Inhibitors
The integration of allosteric inhibition and multivalent targeting represents a paradigm shift in therapeutic approaches to challenging targets like the STAT3 SH2 domain. Computational methods have accelerated the identification of novel allosteric binders from natural compound libraries, with candidates such as ZINC67910988 showing promising binding characteristics and stability [5]. Meanwhile, advances in protein engineering and chemical biology have enabled the rational design of multivalent constructs with enhanced affinity and selectivity [75] [76].
The clinical validation of allosteric targeting approaches continues to advance, as demonstrated by pirmitegravir—an allosteric HIV-1 integrase inhibitor that recently demonstrated proof of concept in clinical trials, establishing the viability of allosteric mechanisms for therapeutic intervention [78]. Similarly, the characterization of allosteric networks in proteins like SHP2 provides a blueprint for understanding and targeting allosteric regulation in STAT3 [74].
Future directions in this field will likely focus on integrating computational predictions with experimental validation through biophysical and cellular assays, developing multivalent protein-drug conjugates with optimized pharmacokinetic properties, and exploring combination strategies that simultaneously target multiple nodes in STAT3 signaling networks. As these innovative approaches mature, they hold significant promise for delivering transformative therapies for STAT3-driven cancers and other pathologies.
Conclusion
The intrinsic flexibility of STAT SH2 domains, once a formidable challenge, is now recognized as a pivotal asset for therapeutic intervention. This synthesis of foundational knowledge, advanced computational methodologies, and rigorous validation frameworks underscores a paradigm shift from static to dynamic drug design. The successful application of molecular dynamics to reveal cryptic pockets and 'induced-active sites' has already yielded promising small-molecule inhibitors with improved drug-like properties. Future directions must focus on extending simulation timescales to capture rare conformational states, integrating machine learning for accelerated dynamics prediction, and exploring the full therapeutic potential of allosteric modulators. Furthermore, understanding how disease-associated mutations alter SH2 domain energy landscapes will open new avenues for personalized medicine. As these dynamic targeting strategies mature, they hold immense promise for developing highly specific, next-generation therapeutics against STAT-driven cancers and immune disorders, ultimately translating molecular insights into clinical breakthroughs.
References
- 1. SH2 Domains: Folding, Binding and Therapeutical Approaches [https://pmc.ncbi.nlm.nih.gov/articles/PMC9783222/]
- 2. Novel STAT 3 small-molecule inhibitors identified by structure -based... [https://scholars.houstonmethodist.org/en/publications/novel-stat3-small-molecule-inhibitors-identified-by-structure-bas]
- 3. Novel STAT3 Small-Molecule Inhibitors Identified by Structure ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8217378/]
- 4. Novel STAT3 small-molecule inhibitors identified by ... [https://www.sciencedirect.com/science/article/abs/pii/S1043661821002218]
- 5. Computational screening and molecular dynamics of natural ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12638358/]
- 6. Identification of the Linker-SH2 Domain of STAT as ... [https://www.sciencedirect.com/science/article/pii/S1535947620338639]
- 7. Beta-sandwich [https://en.wikipedia.org/wiki/Beta-sandwich]
- 8. Screening of potent STAT3-SH2 domain inhibitors from ... [https://link.springer.com/article/10.1007/s11030-022-10490-w]
- 9. Update on Structure and Function of SH2 Domains [https://www.mdpi.com/1422-0067/26/18/9060]
- 10. Structural Implications of STAT3 and STAT5 SH2 Domain ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6895964/]
- 11. The SH2 domains of Stat1 and Stat2 mediate ... - PubMed [https://pubmed.ncbi.nlm.nih.gov/8605877/]
- 12. Mutational Analysis of the STAT6 SH2 Domain [https://www.sciencedirect.com/science/article/pii/S002192581880535X]
- 13. Molecular dynamics simulations of the conformational ... [https://www.sciencedirect.com/science/article/abs/pii/S1093326309001119]
- 14. STAT5B leukemic mutations, altering SH2 tyrosine 665, have ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11999048/]
- 15. Molecular Mechanisms of SH2- and PTB-Domain-Containing ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3839611/]
- 16. Update on Structure and Function of SH2 Domains [https://pmc.ncbi.nlm.nih.gov/articles/PMC12470777/]
- 17. Loops Govern SH2 Domain Specificity by Controlling Access ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6590088/]
- 18. Accurate sequence-to-affinity models for SH2 domains from ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11703206/]
- 19. CoDIAC: A comprehensive approach for interaction analysis ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11291013/]
- 20. Specificity and regulation of phosphotyrosine signaling ... [https://www.sciencedirect.com/science/article/pii/S2590152420300088]
- 21. Review The SH2 domain: versatile signaling module and ... [https://www.sciencedirect.com/science/article/abs/pii/S1570963904002882]
- 22. Phase separation of disease-associated SHP2 mutants ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7572904/]
- 23. Computational screening and molecular dynamics of ... [https://link.springer.com/article/10.1007/s11030-024-11075-5]
- 24. Lipid-Mediated Assembly of Biomolecular Condensates [https://pmc.ncbi.nlm.nih.gov/articles/PMC12467959/]
- 25. SH2-mediated steric occlusion of the C2 domain regulates ... [https://www.sciencedirect.com/science/article/pii/S0021925825026407]
- 26. Protocol for Molecular Dynamics Simulations of Proteins [https://en.bio-protocol.org/en/bpdetail?id=2051&type=0]
- 27. Best Practices for Foundations in Molecular Simulations ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6884151/]
- 28. Novel STAT3 small-molecule inhibitors identified by structure ... [https://pubmed.ncbi.nlm.nih.gov/33932608/]
- 29. Virtual screening, molecular dynamics and structure– ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7441776/]
- 30. Structure-based virtual screening and molecular dynamics ... [https://www.frontiersin.org/journals/veterinary-science/articles/10.3389/fvets.2023.1340126/full]
- 31. Exploring Meta's Open Molecules 2025 (OMol25) & Universal ... [https://www.rowansci.com/blog/exploring-open-molecules-2025]
- 32. A Record-Breaking Dataset to Train AI Models has Launched [https://newscenter.lbl.gov/2025/05/14/computational-chemistry-unlocked-a-record-breaking-dataset-to-train-ai-models-has-launched/]
- 33. In silico simulations of STAT1 and STAT3 inhibitors predict ... [https://www.sciencedirect.com/science/article/abs/pii/S0014299913008224]
- 34. Discovery of potent STAT3 inhibitors using structure-based ... [https://www.frontiersin.org/journals/oncology/articles/10.3389/fonc.2023.1287797/full]
- 35. Identification of New Shikonin Derivatives as Antitumor ... [https://www.nature.com/articles/s41598-017-02671-7]
- 36. Screening of potent STAT3-SH2 domain inhibitors from ... [https://pubmed.ncbi.nlm.nih.gov/35831728/]
- 37. Wordom update 2: A user-friendly program for the analysis ... [https://www.sciencedirect.com/science/article/pii/S2001037023000272]
- 38. Construction of A Non-Redundant Human SH2 Domain ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5172440/]
- 39. SH2 domain regulation by CCD in D170A variant [https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1010794]
- 40. Discriminating between competing models for the allosteric ... [https://www.sciencedirect.com/science/article/pii/S2001037021004633]
- 41. Study on SHP2 Conformational Transition and Structural ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11721961/]
- 42. Binding specificity of SH2 domains: insight from free energy ... [https://pubmed.ncbi.nlm.nih.gov/18767163/]
- 43. SH2db - Documentation [http://sh2db.ttk.hu/documentation]
- 44. Structure and Dynamics of Drk-SH2 Domain and Its Site- ... [https://www.mdpi.com/1422-0067/25/12/6386]
- 45. Conformational heterogeneity of the BTK PHTH domain ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10945472/]
- 46. Conformational heterogeneity of the BTK PHTH domain ... [https://elifesciences.org/reviewed-preprints/89489v1]
- 47. STAT proteins: a kaleidoscope of canonical and non ... [https://jhoonline.biomedcentral.com/articles/10.1186/s13045-021-01214-y]
- 48. Progress in computational methods and mechanistic insights ... [https://pubs.rsc.org/en/content/articlehtml/2025/nr/d4nr05487c]
- 49. Frontier advances in molecular dynamics simulations for the ... [https://pubs.rsc.org/en/content/articlehtml/2025/ra/d5ra00059a?page=search]
- 50. Enabling Scalable AI-Driven Molecular Dynamics ... [https://developer.nvidia.com/blog/enabling-scalable-ai-driven-molecular-dynamics-simulations/]
- 51. Structure and Dynamics of Drk-SH2 Domain and Its Site ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11203457/]
- 52. Novel mechanism of interaction of p85 subunit ... [https://pubmed.ncbi.nlm.nih.gov/15520002/]
- 53. Phosphorylation of disordered proteins tunes local and global ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11628823/]
- 54. Force field (chemistry) [https://en.wikipedia.org/wiki/Force_field_(chemistry)]
- 55. Solvent model [https://en.wikipedia.org/wiki/Solvent_model]
- 56. Design and application of implicit solvent models in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4045398/]
- 57. A Molecular Model for the Differential Activation of STAT 3 and... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0034306]
- 58. Accurate affinity models for SH2 domains from peptide binding ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12521615/]
- 59. Generating and validating renewable affimer protein ... [https://www.nature.com/articles/s41598-024-79357-4]
- 60. SH2 domain [https://en.wikipedia.org/wiki/SH2_domain]
- 61. The JAK-STAT pathway: from structural biology to cytokine ... [https://www.nature.com/articles/s41392-024-01934-w]
- 62. Article SH3-SH2 Domain Orientation in Src Kinases [https://www.sciencedirect.com/science/article/pii/S0969212602007815]
- 63. SH2 Domain Binding: Diverse FLVRs of Partnership [https://www.frontiersin.org/journals/endocrinology/articles/10.3389/fendo.2020.575220/full]
- 64. Molecular Dynamics Simulation Analysis of JAK1 Initial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12387218/]
- 65. Crystal Structure of the Src Family Kinase Hck SH3-SH2 ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2975169/]
- 66. DRUGGING “UNDRUGGABLE” DISEASE-CAUSING PROTEINS [https://pmc.ncbi.nlm.nih.gov/articles/PMC9480546/]
- 67. Defining the Specificity Space of the Human Src Homology ... [https://www.sciencedirect.com/science/article/pii/S1535947620312007]
- 68. Exploring the Effectiveness of Binding Free Energy ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7032235/]
- 69. Thermal shift assay [https://en.wikipedia.org/wiki/Thermal_shift_assay]
- 70. High Throughput Cellular Thermal Shift Assays in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10915787/]
- 71. Allosteric Inhibition: Mechanism, Cooperativity, Examples [https://microbenotes.com/allosteric-inhibition/]
- 72. Allosteric regulation [https://en.wikipedia.org/wiki/Allosteric_regulation]
- 73. Allosteric inhibition explained through conformational ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7720024/]
- 74. Mechanism of activating mutations and allosteric drug ... [https://www.nature.com/articles/s41467-018-06814-w]
- 75. Strategies for targeting cell surface proteins using ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8895325/]
- 76. Multivalent protein-drug conjugates – An emerging strategy ... [https://www.sciencedirect.com/science/article/pii/S0734975023001209]
- 77. PAD Inhibitors: Multiple Approaches Have Striven to Make ... [https://drughunter.com/articles/pad-inhibitors-multiple-approaches-have-striven-to-make-it-to-clinical-developmen]
- 78. Pirmitegravir demonstrates proof of concept in HIV [https://www.eatg.org/hiv-news/idweek-2025-pirmitegravir-demonstrates-proof-of-concept-in-hiv/]